Co-reporter:Leandro F. Vendruscolo, Amanda J. Roberts
Alcohol (May 2014) Volume 48(Issue 3) pp:277-286
Publication Date(Web):May 2014
DOI:10.1016/j.alcohol.2013.08.006
Co-reporter:Melanie M. Hoefer, Ana B. Sanchez, Ricky Maung, Cyrus M. de Rozieres, Irene C. Catalan, Cari C. Dowling, Victoria E. Thaney, Juan Piña-Crespo, Dongxian Zhang, Amanda J. Roberts, Marcus Kaul
Experimental Neurology (January 2015) Volume 263() pp:221-234
Publication Date(Web):1 January 2015
DOI:10.1016/j.expneurol.2014.09.010
•Methamphetamine binge impairs learning and memory of mice in a long-lasting fashion.•Methamphetamine worsens impaired learning and memory in HIV-1 gp120 transgenic mice.•HIV-1 gp120 and methamphetamine injure neuronal dendrites and synapses.•Methamphetamine-exposed HIV-1 gp120tg mice show reduced post-tetanic potentiation.•Methamphetamine and HIV-1 gp120 in combination dysregulate specific synaptic genes.Methamphetamine (METH) abuse is frequent in individuals infected with human immunodeficiency virus type-1 (HIV-1) and is suspected to aggravate HIV-associated neurocognitive disorders (HAND). METH is a psychostimulant that compromises several neurotransmitter systems and HIV proteins trigger neuronal injury but the combined effects of viral infection and METH abuse are incompletely understood. In this study we treated transgenic mice expressing the HIV envelope protein gp120 in the brain (HIV-1 gp120tg) at 3–4 months of age with an escalating-dose, multiple-binge METH regimen. The long-term effects were analyzed after 6–7 months of drug abstinence employing behavioral tests and analysis of neuropathology, electrophysiology and gene expression. Behavioral testing showed that both HIV-1 gp120tg and WT animals treated with METH displayed impaired learning and memory. Neuropathological analysis revealed that METH similar to HIV-1 gp120 caused a significant loss of neuronal dendrites and pre-synaptic terminals in hippocampus and cerebral cortex of WT animals. Electrophysiological studies in hippocampal slices showed that METH exposed HIV-1 gp120tg animals displayed reduced post-tetanic potentiation, whereas both gp120 expression and METH lead to reduced long-term potentiation. A quantitative reverse transcription-polymerase chain reaction array showed that gp120 expression, METH and their combination each caused a significant dysregulation of specific components of GABAergic and glutamatergic neurotransmission systems, providing a possible mechanism for synaptic dysfunction and behavioral impairment. In conclusion, both HIV-1 gp120 and METH caused lasting behavioral impairment in association with neuropathology and altered gene expression. However, combined METH exposure and HIV-1 gp120 expression resulted in the most pronounced, long lasting pre- and post-synaptic alterations coinciding with impaired learning and memory.Download full-size image
Co-reporter:L. Prut, D. Abramowski, T. Krucker, C.L. Levy, A.J. Roberts, M. Staufenbiel, C. Wiessner
Behavioural Brain Research (16 April 2007) Volume 179(Issue 1) pp:
Publication Date(Web):16 April 2007
DOI:10.1016/j.bbr.2007.01.017
Spatial learning and memory deficits in the APP23 transgenic mice have mainly been studied using the Morris water maze (MWM). However learning in the MWM relies on swimming abilities and may be confounded by the stressful nature of this test. We have therefore assessed spatial learning and memory in 12-month-old APP23 using a dry-land maze test developed by Barnes. Mice were given daily learning trials for a total of 41 successive days. After a 12-day interval the mice were re-tested for 4 additional days in order to examine the spatial memory retention. Immediately following this phase, reversal learning was examined for 13 additional days by moving the escape tunnel to the opposite position. During the initial learning phase, APP23 mice showed a significantly longer latency to find the escape tunnel as well as an increased number of errors compared to non-transgenic littermates. These deficits appeared to be due to a delay in switching from a “no strategy” to a spatial strategy. Indeed, this same delay in the use of spatial strategy was observed in the reversal phase of the study. Our results suggest that impairments in APP23 mice in learning and memory maze tests may be due to a specific deficit in the use of spatial strategy.
Co-reporter:Amanda J. Roberts, Ricky Maung, Natalia E. Sejbuk, Christopher Ake, Marcus Kaul
Journal of Neuroscience Methods (15 February 2010) Volume 186(Issue 2) pp:222-225
Publication Date(Web):15 February 2010
DOI:10.1016/j.jneumeth.2009.11.007
The use of drugs for recreational purposes, in particular Methamphetamine, is associated with an increased risk of infection with human immunodeficiency virus (HIV)-1. HIV-1 infection in turn can lead to HIV-associated neurological disorders (HAND) that range from mild cognitive and motor impairment to HIV-associated dementia (HAD). Interestingly, post mortem brain specimens from HAD patients and transgenic (tg) mice expressing the viral envelope protein gp120 in the central nervous system display similar neuropathological signs. In HIV patients, the use of Methamphetamine appears to aggravate neurocognitive alterations. In the present study, we injected HIV/gp120tg mice and non-transgenic littermate control animals with Methamphetamine dissolved in Saline or Saline vehicle and assessed locomotion and stereotyped behaviour. We found that HIVgp120-transgenic mice differ significantly from non-transgenic controls in certain domains of their behavioural response to Methamphetamine. Thus this experimental model system may be useful to further study the mechanistic interaction of both the viral envelope protein and the psychostimulant drug in behavioural alterations and neurodegenerative disease.
Co-reporter:Gor Sarkisyan, Amanda J. Roberts, Peter B. Hedlund
Behavioural Brain Research (1 May 2010) Volume 209(Issue 1) pp:
Publication Date(Web):1 May 2010
DOI:10.1016/j.bbr.2010.01.022
The 5-HT7 receptor has been suggested as a target for treating depression since inactivation or blockade of the receptor has an antidepressant-like behavioral effect. The present study investigated possible interactions between various classes of drugs with antidepressant properties and blockade or inactivation of the 5-HT7 receptor. Immobility despair in the tail suspension test and the forced swim test was evaluated in mice lacking the 5-HT7 receptor (5-HT7−/−) and in wild-type controls (5-HT7+/+) following acute drug treatments. Citalopram, a selective serotonin reuptake inhibitor and widely used antidepressant, dose-dependently reduced immobility in the tail suspension test in both 5-HT7+/+ and 5-HT7−/− mice. Combining doses of citalopram and the 5-HT7 receptor antagonist SB-269970 that by themselves did not affect behavior, reduced immobility in 5-HT7+/+ mice in both the tail suspension test and the forced swim test. No effect was seen in 5-HT7−/− mice. Desipramine and reboxetine, two norepinephrine reuptake inhibitors, dose-dependently reduced immobility in the tail suspension test in 5-HT7+/+ mice, but had no effect in 5-HT7−/− mice. A synergistic effect between desipramine and SB-269970 was found in both behavioral tests in 5-HT7+/+ mice. Reboxetine combined with SB-269970 had effect only in the forced swim test. GBR 12909, a dopamine reuptake inhibitor, dose-dependently reduced tail suspension test immobility in both genotypes. There was no interaction between GBR 12909 and SB-269970. Aripiprazole, an antipsychotic, reduced immobility in both tests in 5-HT7+/+ mice, but not in 5-HT7−/− mice. The results show that the 5-HT7 receptor is required for the observed interaction between this receptor and antidepressants such as citalopram. The data furthermore support the hypothesis that the 5-HT7 receptor might be a suitable target for treating depression.

![4,8:11,15-Dimethano-20H-bisbenzofuro[2,3-a:3',2'-i]dipyrido[4,3-b:3',4'-h]carbazole-1,8a,10a,18-tetrol,7,12-bis(cyclopropylmethyl)-5,6,7,8,9,10,11,12,13,14,19a,20b-dodecahydro-,(4bS,8R,8aS,10aS,11R,14aS,19aR,20bR)-](http://img.cochemist.com/ccimg/105700/105618-26-6.png)
![4,8:11,15-Dimethano-20H-bisbenzofuro[2,3-a:3',2'-i]dipyrido[4,3-b:3',4'-h]carbazole-1,8a,10a,18-tetrol,7,12-bis(cyclopropylmethyl)-5,6,7,8,9,10,11,12,13,14,19a,20b-dodecahydro-,(4bS,8R,8aS,10aS,11R,14aS,19aR,20bR)-](http://img.cochemist.com/ccimg/105700/105618-26-6_b.png)
![AMPEROZIDE HYDROCHLORIDE;4-[4,4-BIS(4-FLUOROPHENYL)BUTYL]-N-ETHYL-1-PIPERAZINECARBOXAMIDEHYDROCHLORIDE](http://img.cochemist.com/ccimg/75600/75558-90-6.png)
![AMPEROZIDE HYDROCHLORIDE;4-[4,4-BIS(4-FLUOROPHENYL)BUTYL]-N-ETHYL-1-PIPERAZINECARBOXAMIDEHYDROCHLORIDE](http://img.cochemist.com/ccimg/75600/75558-90-6_b.png)




















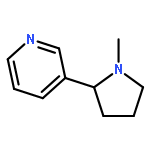
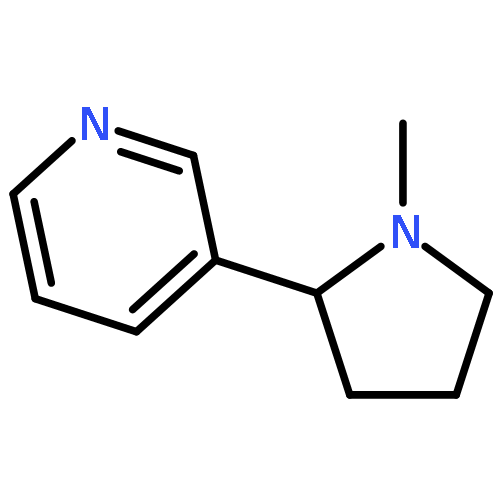


![3-({(2R)-2-[2-(4-methylpiperidin-1-yl)ethyl]pyrrolidin-1-yl}sulfonyl)phenol hydrochloride (1:1)](/data/chemimg/2900/201038-74-6.png)
![3-({(2R)-2-[2-(4-methylpiperidin-1-yl)ethyl]pyrrolidin-1-yl}sulfonyl)phenol hydrochloride (1:1)](/data/chemimg/2900/201038-74-6_b.png)


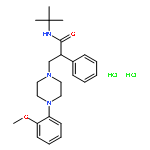
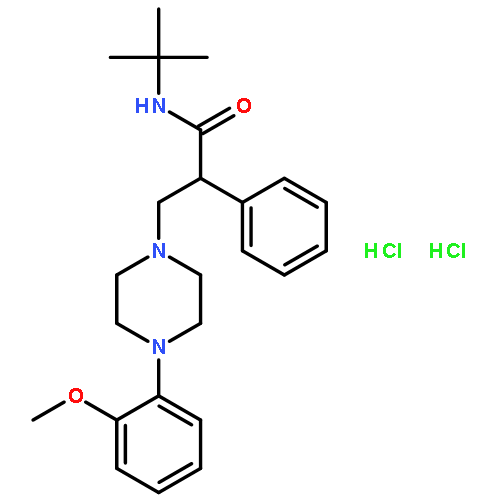

![3-(10,11-Dihydro-5H-dibenzo[b,f]azepin-5-yl)-N-methylpropan-1-amine](http://img.cochemist.com/ccimg/100/50-47-5.png)
![3-(10,11-Dihydro-5H-dibenzo[b,f]azepin-5-yl)-N-methylpropan-1-amine](http://img.cochemist.com/ccimg/100/50-47-5_b.png)
![Methyl (3s,4r)-3-benzoyloxy-8-methyl-8-azabicyclo[3.2.1]octane-4-carboxylate](http://img.cochemist.com/ccimg/100/50-36-2.png)
![Methyl (3s,4r)-3-benzoyloxy-8-methyl-8-azabicyclo[3.2.1]octane-4-carboxylate](http://img.cochemist.com/ccimg/100/50-36-2_b.png)