Co-reporter:Paul R. Carey, Blake R. Gibson, Jordan F. Gibson, Michael E. Greenberg, Hossein Heidari-Torkabadi, Marianne Pusztai-Carey, Sean T. Weaver, and Grant R. Whitmer
Biochemistry May 2, 2017 Volume 56(Issue 17) pp:2247-2247
Publication Date(Web):April 18, 2017
DOI:10.1021/acs.biochem.7b00116
Two protocols that allow for the comparison of Raman spectra of planktonic cells and biofilm formed from these cells in their growth phase have been developed. Planktonic cells are washed and flash-frozen in <1 min to reduce the time for metabolic changes during processing, prior to freeze-drying. Biofilm is formed by standing cells in 50 μL indentations in aluminum foil in an atmosphere of saturated water vapor for 24–48 h. The results for Escherichia coli type K12 cells, which do not readily form biofilm, are compared to those for Staphylococcus epidermidis cells, which prolifically synthesize biofilm. For E. coli, the Raman spectra of the planktonic and biofilm samples are similar with the exception that the spectral signature of RNA, present in planktonic cells, could not be detected in biofilm. For S. epidermidis, major changes occur upon biofilm formation. In addition to the absence of the RNA features, new bands occur near 950 cm–1 and between 1350 and 1420 cm–1 that are associated with an increase in carbohydrate content. Unlike the case in E. coli biofilm, the intensity of G base ring modes is reduced in but A and T base ring signatures become more prominent. For S. epidermis in the biofilm’s amide III region, there is evidence of an increase in the level of β-sheet structure accompanied by a decrease in α-helical content. The presence of biofilm is confirmed by microscope-aided photography and, separately, by staining with methyl violet.
Co-reporter:Hossein Heidari-Torkabadi; Christopher R. Bethel; Zhe Ding; Marianne Pusztai-Carey; Richard Bonnet; Robert A. Bonomo
Journal of the American Chemical Society 2015 Volume 137(Issue 40) pp:12760-12763
Publication Date(Web):September 30, 2015
DOI:10.1021/jacs.5b10007
CTX-M β-lactamases are one of the fastest growing extended-spectrum β-lactamase (ESBL) families found in Escherichia coli rendering this organism extremely difficult to treat with β-lactam antibiotics. Although they are grouped in class A β-lactamases, the CTX-M family possesses low sequence identity with other enzymes. In addition, they have high hydrolytic activity against oxyimino-cephalosporins, despite having smaller active sites compared to other ESBLs in class A. Similar to most class A enzymes, most of the CTX-M β-lactamases can be inhibited by the clinical inhibitors (clavulanic acid, sulbactam, and tazobactam), but the prevalence of inhibitor resistance is an emerging clinical threat. Thus, the mechanistic details of inhibition pathways are needed for new inhibitor development. Here, we use Raman microscopy to study the CTX-M-9 inactivation reaction with the three commercially available inhibitors and compare these findings to the analysis of the S130G variant. Characterization of the reactions in CTX-M-9 single crystals and solution show the formation of a unique cross-linked species, probably involving Ser70 and Ser130, with subsequent hydrolysis leading to an acrylate species linked to Ser130. In solution, a major population of this species is seen at 25 ms after mixing. Support for this finding comes from the CTX-M-9 S130G variant that reacts with clavulanic acid, sulbactam, and tazobactam in solution, but lacks the characteristic spectroscopic signature for the Ser130-linked species. Understanding the mechanism of inactivation of this clinically important ESBL-type class A lactamase permits us to approach the challenge of inhibitor resistance using knowledge of the bridging species in the inactivation pathway.
Co-reporter:Tao Che, Elizabeth A. Rodkey, Christopher R. Bethel, Sivaprakash Shanmugam, Zhe Ding, Marianne Pusztai-Carey, Michael Nottingham, Weirui Chai, John D. Buynak, Robert A. Bonomo, Focco van den Akker, and Paul R. Carey
Biochemistry 2015 Volume 54(Issue 3) pp:734-743
Publication Date(Web):December 23, 2014
DOI:10.1021/bi501197t
For the class A β-lactamase SHV-1, the kinetic and mechanistic properties of the clinically used inhibitor sulbactam are compared with the sulbactam analog substituted in its 6β position by a CH2OH group (6β-(hydroxymethyl)penicillanic acid). The 6β substitution improves both in vitro and microbiological inhibitory properties of sulbactam. Base hydrolysis of both compounds was studied by Raman and NMR spectroscopies and showed that lactam ring opening is followed by fragmentation of the dioxothiazolidine ring leading to formation of the iminium ion within 3 min. The iminium ion slowly loses a proton and converts to cis-enamine (which is a β-aminoacrylate) in 1 h for sulbactam and in 4 h for 6β-(hydroxymethyl) sulbactam. Rapid mix–rapid freeze Raman spectroscopy was used to follow the reactions between the two sulfones and SHV-1. Within 23 ms, a 10-fold excess of sulbactam was entirely hydrolyzed to give a cis-enamine product. In contrast, the 6β-(hydroxymethyl) sulbactam formed longer-lived acyl–enzyme intermediates that are a mixture of imine and enamines. Single crystal Raman studies, soaking in and washing out unreacted substrates, revealed stable populations of imine and trans-enamine acyl enzymes. The corresponding X-ray crystallographic data are consonant with the Raman data and also reveal the role played by the 6β-hydroxymethyl group in retarding hydrolysis of the acyl enzymes. The 6β-hydroxymethyl group sterically hinders approach of the water molecule as well as restraining the side chain of E166 that facilitates hydrolysis.
Co-reporter:Ioanna H. Antonopoulos, Yuko Murayama, Brittany A. Warner, Shun-ichi Sekine, Shigeyuki Yokoyama, and Paul R. Carey
Biochemistry 2015 Volume 54(Issue 3) pp:652-665
Publication Date(Web):December 24, 2014
DOI:10.1021/bi501166r
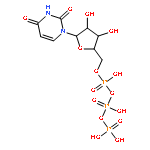
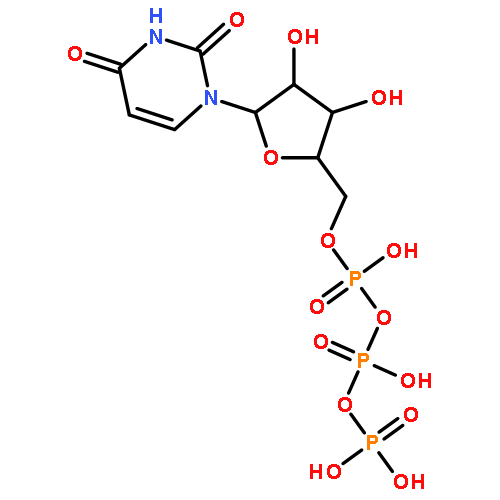
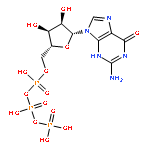
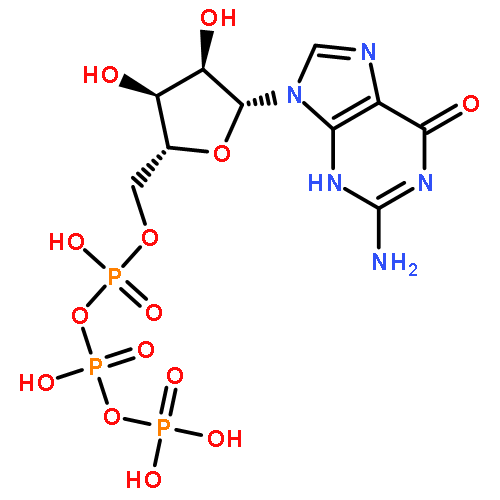
The bacterial RNA polymerase (RNAP) elongation complex (EC) is highly stable and is able to extend an RNA chain for thousands of nucleotides. Understanding the processive mechanism of nucleotide addition requires detailed structural and temporal data for the EC reaction. Here, a time-resolved Raman spectroscopic analysis is combined with polyacrylamide gel electrophoresis (PAGE) to monitor nucleotide addition in single crystals of the Thermus thermophilus EC (TthEC) RNAP. When the cognate base GTP, labeled with 13C and 15N (*GTP), is soaked into crystals of the TthEC, changes in the Raman spectra show evidence of nucleotide incorporation and product formation. The major change is the reduction of *GTP’s triphosphate intensity. Nucleotide incorporation is confirmed by PAGE assays. Both Raman and PAGE methods have a time resolution of minutes. There is also Raman spectroscopic evidence of a second population of *GTP in the crystal that does not become covalently linked to the nascent RNA chain. When this population is removed by “soaking out” (placing the crystal in a solution that contains no NTP), there are no perturbations to the Raman difference spectra, indicating that conformational changes are not detected in the EC. In contrast, the misincorporation of the noncognate base, 13C- and 15N-labeled UTP (*UTP), gives rise to large spectroscopic changes. As in the GTP experiment, reduction of the triphosphate relative intensity in the Raman soak-in data shows that the incorporation reaction occurs during the first few minutes of our instrumental dead time. This is also confirmed by PAGE analysis. Whereas PAGE data show *GTP converts 100% of the nascent RNA 14mer to 15mer, the noncognate *UTP converts only ∼50%. During *UTP soak-in, there is a slow, reversible formation of an α-helical amide I band in the Raman difference spectra peaking at 40 min. Similar to *GTP soak-in, *UTP soak-in shows Raman spectoscopic evidence of a second noncovalently bound *UTP population in the crystal. Moreover, the second population has a marked effect on the complex’s conformational states because removing it by “soaking-out” unreacted *UTP causes large changes in protein and nucleic acid Raman marker bands in the time range of 10–100 min. The conformational changes observed for noncognate *UTP may indicate that the enzyme is preparing for proofreading to excise the misincorporated base. This idea is supported by the PAGE results for *UTP soak-out that show endonuclease activity is occurring.
Co-reporter:Ioanna H. Antonopoulos, Brittany A. Warner, and Paul R. Carey
Biochemistry 2015 Volume 54(Issue 34) pp:
Publication Date(Web):July 29, 2015
DOI:10.1021/acs.biochem.5b00484
Transcription elongation requires the continuous incorporation of ribonucleotide triphosphates into a growing transcript. RNA polymerases (RNAPs) are able to processively synthesize a growing RNA chain via translocation of the RNAP enzyme along its nucleic acid template strand after each nucleotide addition cycle. In this work, a time-resolved Raman spectroscopic analysis of nucleotide addition in single crystals of the Thermus thermophilus elongation complex (TthEC) is reported. When [13C,15N]GTP (*GTP) is soaked into crystals of the TthEC, large reversible changes in the Raman spectrum that are assigned to protein and nucleic acid conformational events during a single-nucleotide incorporation are observed. The *GTP population in the TthEC crystal reaches a stable population at 37 min, while substantial and reversible protein conformational changes (mainly ascribed to changes in α-helical Raman features) maximize at approximately 50 min. At the same time, changes in nucleic acid bases and phosphodiester backbone Raman marker bands occur. Catalysis begins at approximately 65–70 min, soon after the maximal protein and DNA changes, and is monitored via the decline in a triphosphate vibrational Raman mode from *GTP. The Raman data indicate that approximately 40% of the total triphosphate population, present as *GTP, reacts in the crystal. This may suggest that a second population of noncovalently bound *GTP resides in a site distinct from the catalytic site. The data reported here are an extension of our recent work on the elongation complex (EC) of a bacterial RNAP, Thermus thermophilus (Tth), where Raman spectroscopy and polyacrylamide gel electrophoresis were employed to monitor incorporation and misincorporation in single TthEC crystals [Antonopoulos, I. H., et al. (2015) Biochemistry 54, 652–665]. Therefore, the initial study establishes the groundwork for this study. In contrast to our previous study, in which incorporation takes place very rapidly inside the crystals, the data on this single crystal exhibit a slower time regime, which allows the dissection of the structural dynamics associated with GMP incorporation within the TthEC crystal.
Co-reporter:Hossein Heidari-Torkabadi, Tao Che, Michael N. Lombardo, Dennis L. Wright, Amy C. Anderson, and Paul R. Carey
Biochemistry 2015 Volume 54(Issue 17) pp:2719-2726
Publication Date(Web):April 14, 2015
DOI:10.1021/acs.biochem.5b00202
We report the first Raman spectroscopic study of propargyl-linked dihydrofolate reductase (DHFR) inhibitors being taken up by wild type Escherichia coli, Klebsiella pneumoniae, and Staphylococcus aureus cells. A novel protocol is developed where cells are exposed to the fermentation medium containing a known amount of an inhibitor. At a chosen time point, the cells are centrifuged and washed to remove the extracellular compound, then frozen and freeze–dried. Raman difference spectra of the freeze–dried cells (cells exposed to the drug minus cells alone) provide spectra of the compounds inside the cells, where peak intensities allow us to quantify the number of inhibitors within each cell. A time course for the propargyl-linked DHFR inhibitor UCP 1038 soaking into E. coli cells showed that penetration occurs very quickly and reaches a plateau after 10 min exposure to the inhibitor. After 10 min drug exposure, the populations of two inhibitors, UCP 1038 and UCP 1089, were ∼1.5 × 106 molecules in each E. coli cell, ∼4.7 × 105 molecules in each K. pneumonia cell, and ∼2.7 × 106 in each S. aureus cell. This is the first in situ comparison of inhibitor population in Gram-negative and Gram-positive bacterial cells. The positions of the Raman peaks also reveal the protonation of diaminopyrimidine ring upon binding to DHFR inside cells. The spectroscopic signature of protonation was characterized by binding an inhibitor to a single crystal of DHFR.
Co-reporter:Hossein Heidari Torkabadi, Christopher R. Bethel, Krisztina M. Papp-Wallace, Piet A. J. de Boer, Robert A. Bonomo, and Paul R. Carey
Biochemistry 2014 Volume 53(Issue 25) pp:4113-4121
Publication Date(Web):June 5, 2014
DOI:10.1021/bi500529c
Raman microspectroscopy combined with Raman difference spectroscopy reveals the details of chemical reactions within bacterial cells. The method provides direct quantitative data on penetration of druglike molecules into Escherichia coli cells in situ along with the details of drug–target reactions. With this label-free technique, clavulanic acid and tazobactam can be observed as they penetrate into E. coli cells and subsequently inhibit β-lactamase enzymes produced within these cells. When E. coli cells contain a β-lactamase that forms a stable complex with an inhibitor, the Raman signature of the known enamine acyl–enzyme complex is detected. From Raman intensities it is facile to measure semiquantitatively the number of clavulanic acid molecules taken up by the lactamase-free cells during growth.
Co-reporter:Hossein Heidari Torkabadi ; Tao Che ; Jingjing Shou ; Sivaprakash Shanmugam ; Michael W. Crowder ; Robert A. Bonomo ; Marianne Pusztai-Carey
Journal of the American Chemical Society 2013 Volume 135(Issue 8) pp:2895-2898
Publication Date(Web):February 13, 2013
DOI:10.1021/ja311440p
Rapid mix–rapid freeze is a powerful method to study the mechanisms of enzyme–substrate reactions in solution. Here we report a protocol that combines this method with normal (non-resonance) Raman microscopy to enable us to define molecular details of intermediates at early time points. With this combined method, SHV-1, a class A β-lactamase, and tazobactam, a commercially available β-lactamase inhibitor, were rapidly mixed on the millisecond time scale and then were flash-frozen by injection into an isopentane solution surrounded by liquid nitrogen. The “ice” was finally freeze-dried and characterized by Raman microscopy. We found that the reaction is almost complete in solution at 25 ms, giving rise to a major population composed of the trans-enamine intermediate. Between 25 and 500 ms, minor populations of protonated imine are detected that have previously been postulated to precede enamine intermediates. However, within 1 s, the imines are converted entirely to enamines. Interestingly, with this method, we can measure directly the turnover number of SHV-1 and tazobactam. The enzyme is completely inhibited at 1:4 ratio (enzyme:inhibitor) or greater, a number that agrees with the turnover number derived from steady-state kinetic methods. This application, employing non-intensity-enhanced Raman spectroscopy, provides a general and effective route to study the early events in enzyme–substrate reactions.
Co-reporter:Shirly J. Espinoza-Herrera, Vineet Gaur, Zucai Suo, and Paul R. Carey
Biochemistry 2013 Volume 52(Issue 29) pp:
Publication Date(Web):July 3, 2013
DOI:10.1021/bi400524h
Y-Family DNA polymerases are known to bypass DNA lesions in vitro and in vivo. Sulfolobus solfataricus DNA polymerase (Dpo4) was chosen as a model Y-family enzyme for investigating the mechanism of DNA synthesis in single crystals. Crystals of Dpo4 in complexes with DNA (the binary complex) in the presence or absence of an incoming nucleotide were analyzed by Raman microscopy. 13C- and 15N-labeled d*CTP, or unlabeled dCTP, were soaked into the binary crystals with G as the templating base. In the presence of the catalytic metal ions, Mg2+ and Mn2+, nucleotide incorporation was detected by the disappearance of the triphosphate band of dCTP and the retention of *C modes in the crystal following soaking out of noncovalently bound C(or *C)TP. The addition of the second coded base, thymine, was observed by adding cognate dTTP to the crystal following a single d*CTP addition. Adding these two bases caused visible damage to the crystal that was possibly caused by protein and/or DNA conformational change within the crystal. When d*CTP is soaked into the Dpo4 crystal in the absence of Mn2+ or Mg2+, the primer extension reaction did not occur; instead, a ternary protein·template·d*CTP complex was formed. In the Raman difference spectra of both binary and ternary complexes, in addition to the modes of d(*C)CTP, features caused by ring modes from the template/primer bases being perturbed and from the DNA backbone appear, as well as features from perturbed peptide and amino acid side chain modes. These effects are more pronounced in the ternary complex than in the binary complex. Using standardized Raman intensities followed as a function of time, the C(*C)TP population in the crystal was maximal at ∼20 min. These remained unchanged in the ternary complex but declined in the binary complexes as chain incorporation occurred.
Co-reporter:Tao Che ; Robert A. Bonomo ; Sivaprakash Shanmugam ; Christopher R. Bethel ; Marianne Pusztai-Carey ; John D. Buynak
Journal of the American Chemical Society 2012 Volume 134(Issue 27) pp:11206-11215
Publication Date(Web):June 15, 2012
DOI:10.1021/ja303168n
The class D β-lactamases are characterized by the presence of a carboxylated lysine in the active site that participates in catalysis. Found in Acinetobacter baumannii, OXA-24 is a class D carbapenem hydrolyzing enzyme that exhibits resistance to most available β-lactamase inhibitors. In this study, the reaction between a 6-alkylidiene penam sulfone inhibitor, SA-1-204, in single crystals of OXA-24 is followed by Raman microscopy. Details of its reaction with SA-1-204 provide insight into the enzyme’s mode of action and help define the mechanism of inhibition. When the crystal is maintained in HEPES buffer, the reaction is fast, shorter than the time scale of the Raman experiment. However, when the crystal holding solution contains 28% PEG 2000, the reaction is slower and can be recorded by Raman microscopy in real time; the inhibitor’s Raman bands quickly disappear, transient features are seen due to an early intermediate, and, at approximately 2–11 min, new bands appear that are assigned to the late intermediate species. At about 50 min, bands due to all intermediates are replaced by Raman signals of the unreacted inhibitor. The new population remains unchanged indicating (i) that the OXA-24 is no longer active and (ii) that the decarboxylation of Lys84 occurred during the first reaction cycle. Using absorbance spectroscopy, a one-cycle reaction could be carried out in aqueous solution producing inactive OXA-24 as assayed by the chromogenic substrate nitrocefin. However, activity could be restored by reacting aqueous OXA-24 with a large excess of NaHCO3, which recarboxylates Lys84. In contrast, the addition of NaHCO3 was not successful in reactivating OXA-24 in the crystalline state; this is ascribed to the inability to create a concentration of NaHCO3 in large excess over the OXA-24 that is present in the crystal. The finding that inhibitor compounds can inactivate a class D enzyme by promoting decarboxylation of an active site lysine suggests a novel function that could be exploited in inhibitor design.
Co-reporter:Joseph A. Liberman ; Man Guo ; Jermaine L. Jenkins ; Jolanta Krucinska ; Yuanyuan Chen ; Paul R. Carey ;Joseph E. Wedekind
Journal of the American Chemical Society 2012 Volume 134(Issue 41) pp:16933-16936
Publication Date(Web):September 18, 2012
DOI:10.1021/ja3070528
One mechanism by which ribozymes can accelerate biological reactions is by adopting folds that favorably perturb nucleobase ionization. Herein we used Raman crystallography to directly measure pKa values for the Ade38 N1 imino group of a hairpin ribozyme in distinct conformational states. A transition-state analogue gave a pKa value of 6.27 ± 0.05, which agrees strikingly well with values measured by pH–rate analyses. To identify the chemical attributes that contribute to the shifted pKa, we determined crystal structures of hairpin ribozyme variants containing single-atom substitutions at the active site and measured their respective Ade38 N1 pKa values. This approach led to the identification of a single interaction in the transition-state conformation that elevates the base pKa > 0.8 log unit relative to the precatalytic state. The agreement of the microscopic and macroscopic pKa values and the accompanying structural analysis supports a mechanism in which Ade38 N1(H)+ functions as a general acid in phosphodiester bond cleavage. Overall the results quantify the contribution of a single electrostatic interaction to base ionization, which has broad relevance for understanding how RNA structure can control chemical reactivity.
Co-reporter:Yuanyuan Chen ; Ritwika Basu ; Michael L. Gleghorn ; Katsuhiko S. Murakami
Journal of the American Chemical Society 2011 Volume 133(Issue 32) pp:12544-12555
Publication Date(Web):July 11, 2011
DOI:10.1021/ja201557w
The nucleotidyl transfer reaction leading to formation of the first phosphodiester bond has been followed in real time by Raman microscopy, as it proceeds in single crystals of the N4 phage virion RNA polymerase (RNAP). The reaction is initiated by soaking nucleoside triphosphate (NTP) substrates and divalent cations into the RNAP and promoter DNA complex crystal, where the phosphodiester bond formation is completed in about 40 min. This slow reaction allowed us to monitor the changes of the RNAP and DNA conformations as well as bindings of substrate and metal through Raman spectra taken every 5 min. Recently published snapshot X-ray crystal structures along the same reaction pathway assisted the spectroscopic assignments of changes in the enzyme and DNA, while isotopically labeled NTP substrates allowed differentiation of the Raman spectra of bases in substrates and DNA. We observed that substrates are bound at 2–7 min after soaking is commenced, the O-helix completes its conformational change, and binding of both divalent metals required for catalysis in the active site changes the conformation of the ribose triphosphate at position +1. These are followed by a slower decrease of NTP triphosphate groups due to phosphodiester bond formation that reaches completion at about 15 min and even slower complete release of the divalent metals at about 40 min. We have also shown that the O-helix movement can be driven by substrate binding only. The kinetics of the in crystallo nucleotidyl transfer reaction revealed in this study suggest that soaking the substrate and metal into the RNAP–DNA complex crystal for a few minutes generates novel and uncharacterized intermediates for future X-ray and spectroscopic analysis.
Co-reporter:Yuanyuan Chen, Nadukkudy V. Eldho, T. Kwaku Dayie and Paul R. Carey
Biochemistry 2010 Volume 49(Issue 16) pp:
Publication Date(Web):March 12, 2010
DOI:10.1021/bi902117w
Raman difference spectroscopy is used to probe the properties of a 36-nt RNA molecule, “D5”, which lies at the heart of the catalytic apparatus in group II introns. For D5 that has all of its adenine residues labeled with 13C and 15N and utilizing Raman difference spectroscopy, we identify the conformationally sensitive −C−O−P−O−C− stretching modes of the unlabeled bonds adjacent to adenine bases, as well as the adenine ring modes themselves. The phosphodiester modes can be assigned to individual adenine residues based on earlier NMR data. The effect of Mg2+ binding was explored by analyzing the Raman difference spectra for [D5 + Mg2+] minus [D5 no Mg2+], for D5 unlabeled, or D5 labeled with 13C/15N-enriched adenine. In both sets of data we assign differential features to G ring modes perturbed by Mg2+ binding at the N7 position. In the A-labeled spectra we attribute a Raman differential near 1450 cm−1 and changes of intensity at 1296 cm−1 to Mg binding at the N7 position of adenine bases. The A and G bases involved in Mg2+ binding again can be identified using earlier NMR results. For the unlabeled D5, a change in the C−O−P−O−C stretch profile at 811 cm−1 upon magnesium binding is due to a “tightening up” (in the sense of a more rigid molecule with less dynamic interchange among competing ribose conformers) of the D5 structure. For adenine-labeled D5, small changes in the adenine backbone bond signatures in the 810−830 cm−1 region suggest that small conformational changes occur in the tetraloop and bulge regions upon binding of Mg2+. The PO2− stretching vibration, near 1100 cm−1, from the nonbridging phosphate groups, probes the effect of Mg2+−hydrate inner-sphere interactions that cause an upshift. In turn, the upshift is modulated by the presence of monovalent cations since in the presence of Na+ and Li+ the upshift is 23 ± 2 cm−1 while in the presence of K+ and Cs+ it is 13 ± 3 cm−1, a finding that correlates with the differences in hydration radii. These subtle differences in electrostatic interactions may be related to observed variations in catalytic activity. For a reconstructed ribozyme comprising domains 1−3 (D123) connected in cis plus domain 5 (D5) supplied in trans, cleavage of spliced exon substrates in the presence of magnesium and K+ or Cs+ is more efficient than that in the presence of magnesium with Na+ or Li+.
Co-reporter:Matthew Kalp, John D. Buynak and Paul R. Carey
Biochemistry 2009 Volume 48(Issue 43) pp:
Publication Date(Web):September 30, 2009
DOI:10.1021/bi901416t
Mechanism-based inhibitors of class A β-lactamases, such as sulbactam, undergo a complex series of chemical reactions in the enzyme active site. Formation of a trans-enamine acyl-enzyme via a hydrolysis-prone imine is responsible for transient inhibition of the enzyme. Although the imine to enamine tautomerization is crucial to inhibition of the enzyme, there are no experimental data to suggest how this chemical transformation is catalyzed in the active site. In this report, we show that E166 acts as a general base to promote the imine to enamine tautomerization.
Co-reporter:Monica A. Totir, Jooyoung Cha, Akihiro Ishiwata, Benlian Wang, Anjaneyulu Sheri, Vernon E. Anderson, John Buynak, Shahriar Mobashery and Paul R. Carey
Biochemistry 2008 Volume 47(Issue 13) pp:
Publication Date(Web):March 7, 2008
DOI:10.1021/bi702348w
The clinically used inhibitors tazobactam and sulbactam are effective in the inhibition of activity of class A β-lactamases, but not for class D β-lactamases. The two inhibitors exhibit a complex multistep profile for their chemistry of inhibition with class A β-lactamases. To compare the inhibition profiles for class A and D enzymes, the reactions were investigated within OXA-10 β-lactamase (a class D enzyme) crystals using a Raman microscope. The favored reaction pathway appears to be distinctly different from that for class A β-lactamases. In contrast to the case of class A enzymes that favor the formation of a key enamine species, the OXA-10 enzyme forms an α,β-unsaturated acrylate (acid or ester). Quantum mechanical calculations support the likely product as the adduct of Ser115 to the acrylate. Few enamine-like species are formed by sulbactam or tazobactam with this enzyme. Taken together, our results show that the facile conversion of the initial imine, formed upon acylation of the active site Ser67, to the cis- and/or trans-enamine is disfavored. Instead, there is a significant population of the imine that could either experience cross-linking to a second nucleophile (e.g., Ser115) or give rise to the α,β-unsaturated product and permanent inhibition. Alternatively, the imine can undergo hydrolysis to regenerate the catalytically active OXA-10 enzyme. This last process is the dominant one for class D β-lactamases since the enzyme is not effectively inhibited. In contrast to sulbactam and tazobactam, the reactions between oxacillin or 6α-hydroxyisopropylpenicillinate (both substrates) and OXA-10 β-lactamase appear much less complex. These compounds lead to a single acyl−enzyme species, the presence of which was confirmed by Raman and MALDI-TOF experiments.
.jpg)
![4-THIA-1-AZABICYCLO[3.2.0]HEPTANE-2-CARBOXYLIC ACID, 6-(HYDROXYMETHYL)-3,3-DIMETHYL-7-OXO-, 4,4-DIOXIDE, (2S,5R,6R)-](http://img.cochemist.com/ccimg/81400/81324-33-6.png)
![4-THIA-1-AZABICYCLO[3.2.0]HEPTANE-2-CARBOXYLIC ACID, 6-(HYDROXYMETHYL)-3,3-DIMETHYL-7-OXO-, 4,4-DIOXIDE, (2S,5R,6R)-](http://img.cochemist.com/ccimg/81400/81324-33-6_b.png)
















![(2r,3z,5r)-3-(2-hydroxyethylidene)-7-oxo-4-oxa-1-azabicyclo[3.2.0]heptane-2-carboxylic Acid](http://img.cochemist.com/ccimg/58100/58001-44-8.png)
![(2r,3z,5r)-3-(2-hydroxyethylidene)-7-oxo-4-oxa-1-azabicyclo[3.2.0]heptane-2-carboxylic Acid](http://img.cochemist.com/ccimg/58100/58001-44-8_b.png)