Co-reporter:Shengfang Sun, Masanori Sono, Jing Du, and John H. Dawson
Biochemistry 2014 Volume 53(Issue 30) pp:4956-4969
Publication Date(Web):June 27, 2014
DOI:10.1021/bi5002757
The coelomic O2-binding hemoglobin dehaloperoxidase (DHP) from the sea worm Amphitrite ornata is a dual-function heme protein that also possesses a peroxidase activity. Two different starting oxidation states are required for reversible O2 binding (ferrous) and peroxidase (ferric) activity, bringing into question how DHP manages the two functions. In our previous study, the copresence of substrate 2,4,6-trichlorophenol (TCP) and H2O2 was found to be essential for the conversion of oxy-DHP to enzymatically active ferric DHP. On the basis of that study, a functional switching mechanism involving substrate radicals (TCP•) was proposed. To further support this mechanism, herein we report details of our investigations into the H2O2-mediated conversion of oxy-DHP to the ferric or ferryl ([TCP] < [H2O2]) state triggered by both biologically relevant [TCP and 4-bromophenol (4-BP)] and nonrelevant (ferrocyanide) compounds. At <50 μM H2O2, all of these conversion reactions are completely inhibited by ferric heme ligands (KCN and imidazole), indicating the involvement of ferric DHP. Furthermore, the spin-trapping reagent 5,5-dimethyl-1-pyrroline-N-oxide (DMPO) effectively inhibits the TCP/4-BP (but not ferrocyanide)-triggered conversion of oxy-DHP to ferric DHP. These results and O2 concentration-dependent conversion rates observed in this study demonstrate that substrate TCP triggers the conversion of oxy-DHP to a peroxidase by TCP• oxidation of the deoxyferrous state. TCP• is progressively generated, by increasingly produced amounts of ferric DHP, upon H2O2 oxidation of TCP catalyzed initially by trace amounts of ferric enzyme present in the oxy-DHP sample. The data presented herein further address the mechanism of how the halophenolic substrate triggers the conversion of hemoglobin DHP into a peroxidase.
Co-reporter:Chunxue Wang, Leslie L. Lovelace, Shengfang Sun, John H. Dawson, and Lukasz Lebioda
Biochemistry 2013 Volume 52(Issue 36) pp:6203-6210
Publication Date(Web):August 16, 2013
DOI:10.1021/bi400627w
The hemoglobin of sea worm Amphitrite ornata, which for historical reasons is abbreviated as DHP for dehaloperoxidase, has two physiological functions: it binds dioxygen in the ferrous state and dehalogenates halophenols, such as 2,4,6-trichlorophenol (TCP), using hydrogen peroxide as the oxidant in the ferric state. The crystal structures of three DHP variants (Y34N, Y34N/S91G, and L100F) with TCP bound show two mutually exclusive modes of substrate binding. One of them, the internal site, is deep inside the distal pocket with the phenolic OH moiety forming a hydrogen bond to the water molecule coordinated to the heme Fe. In this complex, the distal histidine is predominantly located in the closed position and also forms a hydrogen bond to the phenolic hydroxide. The second mode of TCP binding is external, at the heme edge, with the halophenol molecule forming a lid covering the entrance to the distal cavity. The distal histidine is in the open position and forms a hydrogen bond to the OH group of TCP, which also hydrogen bonds to the hydroxyl of Tyr38. The distance between the Cl4 atom of TCP and the heme Fe is 3.9 Å (nonbonding). In both complexes, TCP molecules prevent the approach of hydrogen peroxide to the heme, indicating that the complexes are inhibitory and implying that the substrates must bind in an ordered fashion: hydrogen peroxide first and TCP second. Kinetic studies confirmed the inhibition of DHP by high concentrations of TCP. The external binding mode may resemble the interaction of TCP with Compound I, the catalytic intermediate to which halophenols bind. The measured values of the apparent Km for TCP were in the range of 0.3–0.8 mM, much lower than the concentrations required to observe TCP binding in crystals. This indicates that during catalysis TCP binds to Compound I. Mutant F21W, which likely has the internal TCP binding site blocked, has ∼7% of the activity of wild-type DHP.
Co-reporter:Suganya Sumithran, Masanori Sono, Gregory M. Raner, John H. Dawson
Journal of Inorganic Biochemistry 2012 Volume 117() pp:316-321
Publication Date(Web):December 2012
DOI:10.1016/j.jinorgbio.2012.09.017
Horseradish peroxidase (HRP) catalyzes the oxidative para-dechlorination of the environmental pollutant/carcinogen 2,4,6-trichlorophenol (2,4,6-TCP). A possible mechanism for this reaction is a direct oxygen atom transfer from HRP compound I (HRP I) to trichlorophenol to generate 2,6-dichloro 1,4-benzoquinone, a two-electron transfer process. An alternative mechanism involves two consecutive one-electron transfer steps in which HRP I is reduced to compound II (HRP II) and then to the ferric enzyme as first proposed by Wiese et al. [F.W. Wiese, H.C. Chang, R.V. Lloyd, J.P. Freeman, V.M. Samokyszyn, Arch. Environ. Contam. Toxicol. 34 (1998) 217–222]. To probe the mechanism of oxidative halophenol dehalogenation, the reactions between 2,4,6-TCP and HRP compounds I or II have been investigated under single turnover conditions (i.e., without excess H2O2) using rapid scan stopped-flow spectroscopy. Addition of 2,4,6-TCP to HRP I leads rapidly to HRP II and then more slowly to the ferric resting state, consistent with a mechanism involving two consecutive one-electron oxidations of the substrate via a phenoxy radical intermediate. HRP II can also directly dechlorinate 2,4,6-TCP as judged by rapid scan stopped-flow and mass spectrometry. This observation is particularly significant since HRP II can only carry out one-electron oxidations. A more detailed understanding of the mechanism of oxidative halophenol dehalogenation will facilitate the use of HRP as a halophenol bioremediation catalyst.Single turnover experiments reveal that horseradish peroxidase (HRP) catalyzes the oxidative dehalogenation of halophenols by a mechanism involving two consecutive one-electron oxidations of the substrate via a phenoxy radical intermediate. Further development of HRP as a halophenol bioremediation catalyst will benefit from the more detailed understanding of this mechanism described in this paper.Highlights► The mechanism of halophenol dehalogenation by horseradish peroxidase is examined. ► Stopped-flow experiments reveal both Compounds I and II to be active catalysts. ► Single turnover data establish a mechanism involving two one-electron steps. ► Detailed mechanistic information will foster use of HRP as a bioremediation catalyst.
Co-reporter:Shengfang Sun, Masanori Sono, Chunxue Wang, Jing Du, Lukasz Lebioda, John H. Dawson
Archives of Biochemistry and Biophysics (1 March 2014) Volume 545() pp:108-115
Publication Date(Web):1 March 2014
DOI:10.1016/j.abb.2014.01.010



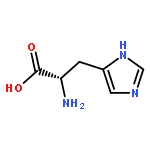
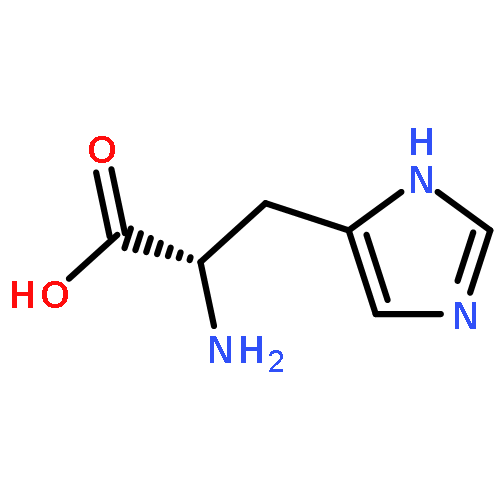







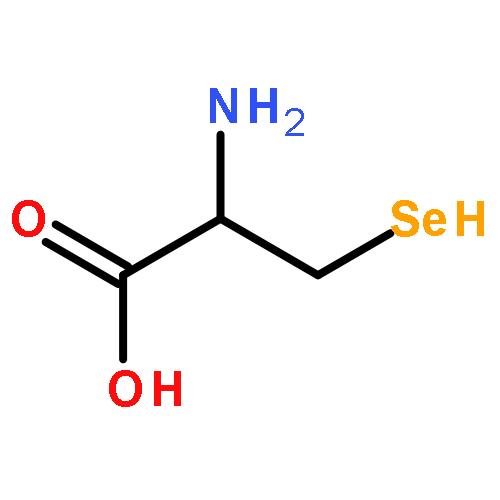
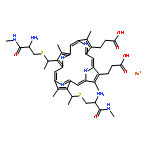







![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8.png)
![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8_b.png)