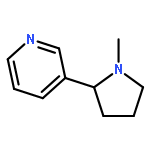
Co-reporter:Olivier George, Bruce T. Hope
Neuropharmacology 2017 Volume 122(Volume 122) pp:
Publication Date(Web):1 August 2017
DOI:10.1016/j.neuropharm.2017.04.031
•Novel approaches to study the neuronal ensembles of alcoholism.•A neuronal ensemble in the infralimbic cortex prevents relapse in dependent rats.•A neuronal ensemble in the central amygdala promotes drinking in dependent rats.•Remaining questions on the timing and consolidation of ensembles are highlighted.•A hypothesis on how these neuronal ensembles may interact is proposed.Alcohol induces many alterations in the brain that are thought to contribute to alcohol addiction. Most of the known alterations are induced in all neurons of a brain area or all neurons of a given cell type, regardless of whether they were activated during behavior. While these alterations can have important modulatory effects on behavior, they cannot explain why animals respond specifically to alcohol-paired cues as opposed to all other non-paired cues and evoke highly specific goal-directed learned responses in models of drug craving. As an alternative, we hypothesize another class of alterations that are induced only within sparsely distributed patterns of neurons, called neuronal ensembles, that are selectively activated by alcohol-specific cues during behavior and encode the long-term memories underlying these learned behaviors in animal models of alcohol addiction. Here we review recent studies and techniques used to identify the role of neuronal ensembles in animal models of different phases of the alcohol addiction cycle.This article is part of the Special Issue entitled “Alcoholism”.
Co-reporter:Giordano de Guglielmo;Marsida Kallupi;Maury D. Cole
Psychopharmacology 2017 Volume 234( Issue 13) pp:2009-2018
Publication Date(Web):24 March 2017
DOI:10.1007/s00213-017-4608-7
A major issue in the addiction field is the limited number of animal models of the voluntary induction and maintenance of alcohol dependence in outbred rats.To address this issue, we developed a novel apparatus that vaporizes alcohol for 2–10 min after an active nosepoke response.Male Wistar rats were allowed to self-administer alcohol vapor for 8 h/day every other day for 24 sessions (escalated) or eight sessions (non-escalated). Escalated and non-escalated rats were then tested for progressive ratio responding. Anxiety-like behavior, somatic signs of withdrawal, and hyperalgesia were assessed during acute withdrawal.The results showed that rats exhibited excellent discrimination between the active and inactive operanda (>85%), and the escalated rats quickly increased their blood alcohol levels from ~50 to >200 mg% in ~6 weeks. Compared with non-escalated rats, escalated rats exhibited severe addiction-like behavior, including somatic signs of withdrawal, anxiety-like behavior, hyperalgesia, and higher responding on a progressive ratio schedule of reinforcement.These results demonstrate that outbred rats will voluntarily self-administer alcohol vapor to the point of dependence without the use of forced alcohol administration, sweeteners, food/water restriction, operant pretraining, or behavioral/genetic selection. This novel animal model may be particularly useful for medication development to help unveil the neuronal circuitry that underlies the voluntary induction of alcohol addiction and identify novel molecular targets that are specifically recruited after the voluntary induction and maintenance of alcohol dependence.
Co-reporter:Olivier George;George F. Koob;Leandro F. Vendruscolo
Psychopharmacology 2014 Volume 231( Issue 19) pp:3911-3917
Publication Date(Web):2014 October
DOI:10.1007/s00213-014-3623-1
Co-reporter:Ami Cohen, George F Koob and Olivier George
Neuropsychopharmacology 2012 37(9) pp:2153-2160
Publication Date(Web):May 2, 2012
DOI:10.1038/npp.2012.67
Although established smokers have a very regular pattern of smoking behavior, converging lines of evidence suggest that the escalation of smoking behavior is a critical factor in the development of dependence. However, the neurobiological mechanisms that underlie the escalation of smoking are unknown, because there is no animal model of the escalation of nicotine intake. On the basis of the pattern of smoking behavior in humans and presence of monoamine oxidase inhibitors in tobacco smoke, we hypothesized that the escalation of nicotine intake may only occur when animals are given extended-access (21 h per day) self-administration sessions after repeated periods of abstinence (24–48 h), and after chronic inhibition of monoamine oxidase using phenelzine sulfate. Intermittent access (every 24–48 h) to extended nicotine self-administration produced a robust escalation of nicotine intake, associated with increased responding under fixed- and progressive-ratio schedules of reinforcement, and increased somatic signs of withdrawal. The escalation of nicotine intake was not observed in rats with intermittent access to limited (1 h per day) nicotine self-administration or daily access to extended (21 h per day) nicotine self-administration. Moreover, inhibition of monoamine oxidase with daily administration of phenelzine increased nicotine intake by ~50%. These results demonstrate that the escalation of nicotine intake only occurs in animals given intermittent periods of abstinence with extended access to nicotine, and that inhibition of monoamine oxidase may contribute to the escalation of smoking, thus validating both an animal model of the escalation of smoking behavior and the contribution of monoamine oxidase inhibition to compulsive nicotine-seeking.
Co-reporter:Chelsea Sanders;John Freiling;Shayla Vu;Edward Grigoryan;Camryn D. Allen;Elena Crawford;Chitra D. Mandyam;George F. Koob
PNAS 2012 Volume 109 (Issue 44 ) pp:18156-18161
Publication Date(Web):2012-10-30
DOI:10.1073/pnas.1116523109
Chronic intermittent access to alcohol leads to the escalation of alcohol intake, similar to binge drinking in humans. Converging
lines of evidence suggest that impairment of medial prefrontal cortex (mPFC) cognitive function and overactivation of the
central nucleus of the amygdala (CeA) are key factors that lead to excessive drinking in dependence. However, the role of
the mPFC and CeA in the escalation of alcohol intake in rats with a history of binge drinking without dependence is currently
unknown. To address this issue, we examined FBJ murine osteosarcoma viral oncogene homolog (Fos) expression in the mPFC, CeA,
hippocampus, and nucleus accumbens and evaluated working memory and anxiety-like behavior in rats given continuous (24 h/d
for 7 d/wk) or intermittent (3 d/wk) access to alcohol (20% vol/vol) using a two-bottle choice paradigm. The results showed
that abstinence from alcohol in rats with a history of escalation of alcohol intake specifically recruited GABA and corticotropin-releasing
factor (CRF) neurons in the mPFC and produced working memory impairments associated with excessive alcohol drinking during
acute (24–72 h) but not protracted (16 –68 d) abstinence. Moreover, abstinence from alcohol was associated with a functional
disconnection of the mPFC and CeA but not mPFC and nucleus accumbens. These results show that recruitment of a subset of GABA
and CRF neurons in the mPFC during withdrawal and disconnection of the PFC–CeA pathway may be critical for impaired executive
control over motivated behavior, suggesting that dysregulation of mPFC interneurons may be an early index of neuroadaptation
in alcohol dependence.
Co-reporter:Olivier George;Allison Lloyd;F. Ivy Carroll;M. Imad Damaj
Psychopharmacology 2011 Volume 213( Issue 4) pp:715-722
Publication Date(Web):2011 February
DOI:10.1007/s00213-010-2024-3
Much evidence indicates that individuals use tobacco primarily to experience the psychopharmacological properties of nicotine. Varenicline, a partial α4β2 nicotinic acetylcholine receptor (nAChR) agonist, is effective in reducing nicotine craving and relapse in smokers, suggesting that α4β2 nAChRs may play a key role in nicotine dependence. In rats, the effect of varenicline on nicotine intake has only been studied with limited access to the drug, a model of the positive reinforcing effect of nicotine. Varenicline has not been tested on the increase in motivation to take nicotine in nicotine-dependent rats.The present study evaluated the effects of varenicline on nicotine intake in rats with extended access to nicotine self-administration (23 h/day), a condition leading to the development of nicotine dependence. We hypothesized that varenicline’s effects on nicotine self-administration would be greater in rats with extended than limited access to the drug and after forced abstinence rather than during baseline self-administration.Varenicline dose-dependently decreased nicotine self-administration in rats with limited (1 h/day) and extended (23 h/day) access. Despite an increased sensitivity to the motivational effects of abstinence on nicotine intake compared with limited-access rats, varenicline was equally effective in decreasing nicotine intake in dependent rats with extended access to nicotine.These results suggest that α4β2 nAChRs are critical in mediating the positive reinforcing effects of nicotine but may not be a key element underlying the negative reinforcement process responsible for the increased nicotine intake after abstinence in dependent subjects.
Co-reporter:Olivier George, Chitra D Mandyam, Sunmee Wee and George F Koob
Neuropsychopharmacology 2008 33(10) pp:2474-2482
Publication Date(Web):November 21, 2007
DOI:10.1038/sj.npp.1301626
Humans with drug addiction exhibit compulsive drug-seeking associated with impairment of prefrontal cortex cognitive function. Whether prefrontal cortex dysfunction is a consequence of chronic drug exposure, or mediates the transition from drug use to drug dependence, is unknown. The current study investigates whether a history of escalated vs controlled cocaine intake is associated with specific working memory impairments, and long-lasting alterations of the dorsomedial prefrontal cortex and orbitofrontal cortex in rats. Working memory was assessed in rats with a history of extended (6 h per session) or limited (1 h per session) access to cocaine (0.5 mg/kg per injection), 3–17 days after the last self-administration session, using a delayed nonmatching-to-sample task. The density of neurons, oligodendrocytes, and astrocytes was quantified in the dorsomedial prefrontal cortex and orbitofrontal prefrontal cortex 2 months after the last self-administration session. Working memory impairments were observed after a history of chronic and escalated cocaine intake, but not after repeated limited access to cocaine. Moreover, working memory impairments were correlated with a decreased density of neurons and oligodendrocytes but not astrocytes in the dorsomedial prefrontal cortex, and with a decreased density of oligodendrocytes in the orbitofrontal cortex. Considering the role of the prefrontal cortex in goal-directed behavior, the prefrontal cortex dysfunctions observed here may exacerbate the loss of control associated with increased drug use and facilitate the progression to drug addiction.
Co-reporter:Pietro Cottone;Eric P. Zorrilla;Loren H. Parsons;Sandy Ghozland;Marc R. Azar;Laura E. O'Dell;Heather N. Richardson;George F. Koob
PNAS 2007 Volume 104 (Issue 43 ) pp:17198-17203
Publication Date(Web):2007-10-23
DOI:10.1073/pnas.0707585104
Nicotine, the main psychoactive ingredient of tobacco, induces negative emotional symptoms during abstinence that contribute
to a profound craving for nicotine. However, the neurobiological mechanisms underlying how nicotine produces dependence remains
poorly understood. We demonstrate one mechanism for both the anxiety-like symptoms of withdrawal and excessive nicotine intake
observed after abstinence, through recruitment of the extrahypothalamic stress peptide corticotropin-releasing factor (CRF)
system and activation of CRF1 receptors. Overactivation of the CRF–CRF1 system may contribute to nicotine dependence and may represent a prominent target for investigating the vulnerability to
tobacco addiction.
Co-reporter:Olivier George, George F. Koob
Neuroscience & Biobehavioral Reviews (November 2010) Volume 35(Issue 2) pp:232-247
Publication Date(Web):1 November 2010
DOI:10.1016/j.neubiorev.2010.05.002
Several neuropsychological hypotheses have been formulated to explain the transition to addiction, including hedonic allostasis, incentive salience, and the development of habits. A key feature of addiction that remains to be explored is the important individual variability observed in the propensity to self-administer drugs, the sensitivity to drug-associated cues, the severity of the withdrawal state, and the ability to quit. In this review, we suggest that the concept of self-regulation, combined with the concept of modularity of cognitive function, may aid in the understanding of the neural basis of individual differences in the vulnerability to drugs and the transition to addiction. The thesis of this review is that drug addiction involves a failure of the different subcomponents of the executive systems controlling key cognitive modules that process reward, pain, stress, emotion, habits, and decision-making. A subhypothesis is that the different patterns of drug addiction and individual differences in the transition to addiction may emerge from differential vulnerability in one or more of the subcomponents.

![4,8:11,15-Dimethano-20H-bisbenzofuro[2,3-a:3',2'-i]dipyrido[4,3-b:3',4'-h]carbazole-1,8a,10a,18-tetrol,7,12-bis(cyclopropylmethyl)-5,6,7,8,9,10,11,12,13,14,19a,20b-dodecahydro-,(4bS,8R,8aS,10aS,11R,14aS,19aR,20bR)-](http://img.cochemist.com/ccimg/105700/105618-26-6.png)
![4,8:11,15-Dimethano-20H-bisbenzofuro[2,3-a:3',2'-i]dipyrido[4,3-b:3',4'-h]carbazole-1,8a,10a,18-tetrol,7,12-bis(cyclopropylmethyl)-5,6,7,8,9,10,11,12,13,14,19a,20b-dodecahydro-,(4bS,8R,8aS,10aS,11R,14aS,19aR,20bR)-](http://img.cochemist.com/ccimg/105700/105618-26-6_b.png)



![N,2,2,3-tetramethylbicyclo[2.2.1]heptan-3-amine](http://img.cochemist.com/ccimg/100/60-40-2.png)
![N,2,2,3-tetramethylbicyclo[2.2.1]heptan-3-amine](http://img.cochemist.com/ccimg/100/60-40-2_b.png)


![Methyl (3s,4r)-3-benzoyloxy-8-methyl-8-azabicyclo[3.2.1]octane-4-carboxylate](http://img.cochemist.com/ccimg/100/50-36-2.png)
![Methyl (3s,4r)-3-benzoyloxy-8-methyl-8-azabicyclo[3.2.1]octane-4-carboxylate](http://img.cochemist.com/ccimg/100/50-36-2_b.png)