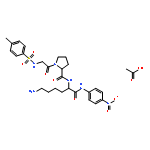
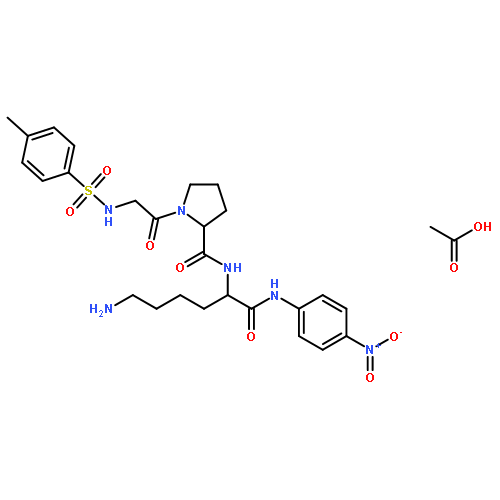
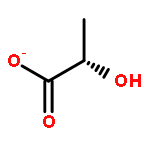
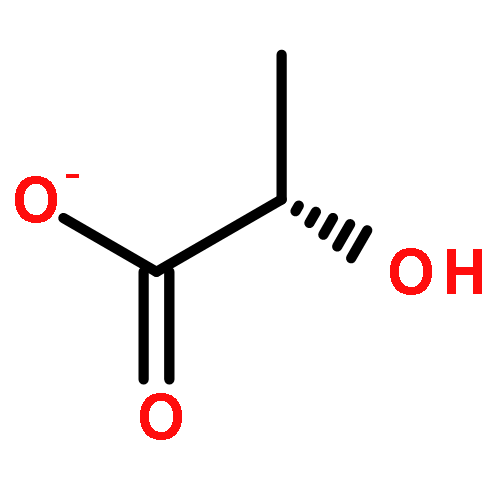
Co-reporter:Shang Gao;Xue Wang;Shasha Wang;Shanshan Zhu;Rui Rong;Xiaolong Xu
Metallomics (2009-Present) 2017 vol. 9(Issue 6) pp:706-714
Publication Date(Web):2017/06/21
DOI:10.1039/C7MT00024C
The wide range of applications of ZnO nanoparticles (nano ZnO) in commercial products and the ubiquitous cadmium (Cd) contamination in the natural environment increase the chance of co-existence of nano ZnO with Cd in the surroundings. To investigate the effects of nano ZnO on CdCl2-induced hepatotoxicity in mice, the histopathologic changes, metallothionein expression, oxidative stress responses and serum biochemical parameters were determined after oral administration of bulk or nano ZnO and/or CdCl2 for seven consecutive days. Bulk or nano ZnO had low toxicity in mice. In contrast, CdCl2 led to significant hepatic oxidative damage, as indicated by hepatic histopathological abnormalities and dysfunction. Bulk and nano ZnO had nearly identical influences on the hepatotoxicity of CdCl2 in mice. Although co-administration of bulk or nano ZnO with CdCl2 had a positive cooperative effect on the hepatic uptake of Cd and Zn, both bulk and nano ZnO significantly attenuated CdCl2-caused hepatic damage via the reduction of oxidative stress. The increase in metallothionein synthesis and the reduction of Cd-induced perturbation of Zn2+ homeostasis after co-administration of bulk or nano ZnO with CdCl2 play two important roles in the protective effect of bulk or nano ZnO on CdCl2-caused hepatic oxidative damage.
Co-reporter:Yan Zhang, Xiaolong Xu, Shanshan Zhu, Jiajia Song, Xincheng Yan and Shang Gao
Toxicology Research 2016 vol. 5(Issue 5) pp:1309-1317
Publication Date(Web):21 Jul 2016
DOI:10.1039/C6TX00190D
Nanomaterials have been widely used in diverse areas. Heavy metals are ubiquitous environmental pollutants. In spite of the real risk of humans’ co-exposure to nanoparticles and heavy metals, their combined toxicity has received little attention. We have reported that silica nanoparticles and CdCl2 have a positive synergistic toxicity in mice. Here, we demonstrate that Fe3O4 nanoparticles (nano-Fe3O4) and CdCl2 have a negative synergistic toxicity in mice. Nano-Fe3O4 showed low toxicity in mice. In contrast, CdCl2 caused significant oxidative damage mainly in the liver as indicated by severe liver dysfunction and histopathological abnormalities. Co-exposure to nano-Fe3O4 and CdCl2 significantly attenuated CdCl2-induced damage in the liver through reduction of oxidative stress. Nano-Fe3O4 and CdCl2 had negative cooperative effects on the biodistributions of Fe and Cd in mice due to mutually competitive inhibition of Fe and Cd uptake. The reduction of Cd accumulation in tissues and the inhibition of Cd-induced deprivation of tissue Fe by nano-Fe3O4 played two key roles in the protective effect of nano-Fe3O4 on CdCl2-induced oxidative damage.
Co-reporter:Xincheng Yan, Rui Rong, Shanshan Zhu, Mingchun Guo, Shang Gao, Shasha Wang, and Xiaolong Xu
Journal of Agricultural and Food Chemistry 2015 Volume 63(Issue 37) pp:8292-8298
Publication Date(Web):September 3, 2015
DOI:10.1021/acs.jafc.5b01979
The extensive applications of ZnO nanoparticles (nano ZnO) and dimethoate have increased the risk of people’s coexposure to nano ZnO and dimethoate. Therefore, we evaluated in this study the effects of nano or bulk ZnO on dimethoate-induced toxicity in mice. The serum biochemical parameters, biodistributions, oxidative stress responses, and histopathological changes in mice were measured after intragastric administration of nano or bulk ZnO and/or dimethoate for 14 days. Oral administration of nano or bulk ZnO at a dose of 50 mg/kg did not cause obvious injury in mice. In contrast, oral administration of dimethoate at a dose of 15 mg/kg induced observable oxidative damage in mice. Co-administration of nano or bulk ZnO with dimethoate significantly increased Zn accumulation by 30.7 ± 1.7% or 29.7 ± 2.4% and dimethoate accumulation by 42.8 ± 2.1% or 46.6 ± 2.9% in the liver, respectively. The increased accumulations of dimethoate and Zn in the liver reduced its cholinesterase activity from 5.64 ± 0.45 U/mg protein to 4.67 ± 0.42 U/mg protein or 4.76 ± 0.45 U/mg protein for nano or bulk ZnO, respectively. Furthermore, the accumulations of dimethoate and Zn in liver also increased hepatic oxidative stress, resulting in severe liver damage. Both nano and bulk ZnO dissolved quickly in acidic gastric fluid, regardless of particle size; therefore, they had nearly identical enhanced effects on dimethoate-induced toxicity in mice.
Co-reporter:Lili Peng, Xiaolong Xu, Mingchun Guo, Xincheng Yan, Shasha Wang, Shang Gao and Shanshan Zhu
Metallomics 2013 vol. 5(Issue 7) pp:920-927
Publication Date(Web):11 Jun 2013
DOI:10.1039/C3MT00031A
Obviously different from the other known phosphodiesterases, the phosphodiesterase from Trimeresurus stejnegeri venom (TS-PDE) consists of two different chains linked with disulfide bonds and contains both endogenous Cu2+ and Zn2+. Cu2+ and Zn2+ are important for its phosphodiesterase activity. In this study, the effects of metal ions and small-molecule reductants on its structure and activity have been investigated by polyacrylamide gel electrophoresis, high performance liquid chromatography, fluorescence and electron paramagnetic resonance spectroscopy. The results show that TS-PDE has one class of Zn2+ binding site and two classes of Cu2+ binding site, including the high affinity activator sites and the low affinity sites. Cu2+ ions function as a switch for its phosphodiesterase activity. The catalytic activity of TS-PDE does not have an absolute requirement for Cu2+ and Zn2+. Mg2+, Mn2+, Ni2+, Co2+ and Ca2+ are all effective for its phosphodiesterase activity. TS-PDE has seven disulfide bonds and ten free cysteine residues. L-Ascorbate inhibits the phosphodiesterase activity of TS-PDE through reduction of the Cu2+, while dithiothreitol, glutathione and tris(2-carboxyethyl)phosphine inhibit the phosphodiesterase activity of TS-PDE by reducing both the Cu2+ and disulfide bonds. The catalytic activity of TS-PDE relies on its disulfide bonds and bimetallic cluster. In addition, biologically-relevant reductants, glutathione and L-ascorbate, have been found to be endogenous inhibitors to the phosphodiesterase activity of TS-PDE.
Co-reporter:Liyun Zhang, Xiaolong Xu, Zhaofeng Luo, Yan Zhang, Dengke Shen, Lili Peng and Jiajia Song
Metallomics 2012 vol. 4(Issue 2) pp:166-173
Publication Date(Web):02 Nov 2011
DOI:10.1039/C1MT00135C
NAD-glycohydrolase (AA-NADase) from Agkistrodon acutus venom is a unique multicatalytic enzyme with both NADase and AT(D)Pase-like activities. Among all identified NADases, only AA-NADase is a disulfide-linked dimer and contains Cu2+. Cu2+ and disulfide bonds are essential for its multicatalytic activity. In this study, the effects of Cu2+ and disulfide-bonds on guanidine hydrochloride (GdnHCl)- and thermal-induced unfolding of AA-NADase have been investigated by fluorescence, circular dichroism (CD) and differential scanning calorimetry (DSC). Cu2+ and disulfide bonds not only increase the free energy change during the GdnHCl-induced unfolding as determined by fluorescence, but also increase the overall enthalpy change and the transition temperature during the thermal-induced unfolding as determined by CD and DSC. The slope of the GdnHCl-induced unfolding curve at its midpoint and the heat capacity of thermal-induced unfolding are slightly affected by Cu2+ but significantly decrease after reduction of three disulfide-bonds. This work suggests that Cu2+ stabilizes the folded state by increasing the enthalpy of unfolding, while disulfide-bonds stabilize the folded state by increasing the enthalpy of unfolding and stabilizing the packing of hydrophobic residues. Thus both Cu2+ and disulfide bonds play a structural role in its multicatalytic activity.
Co-reporter:Deng-Ke Shen;Yan Zhang;Jia-Jia Song;Xin-Cheng Yan ;Ming-Chun Guo
Biopolymers 2012 Volume 97( Issue 10) pp:818-824
Publication Date(Web):
DOI:10.1002/bip.22078
Abstract
Anticoagulation factor II (ACF II), a coagulation factor X- binding protein from the venom of Agkistrodon acutus has both anticoagulant and hypotensive activities. Previous studies show that ACF II binds specifically with activated factor X (FXa) in a Ca2+-dependent manner and inhibits intrinsic coagulation pathway. In this study, the inhibition of extrinsic coagulation pathway by ACF II was measured in vivo by prothrombin time assay and the binding of ACF II to factor IX (FIX) was investigated by native polyacrylamide gel electrophoresis and surface plasmon resonance (SPR). The results indicate that ACF II also inhibits extrinsic coagulation pathway, but does not inhibit thrombin activity. ACF II also binds with FIX with high binding affinity in a Ca2+-dependent manner and their maximal binding occurs at about 0.1 mM Ca2+. ACF II has similar binding affinity to FIX and FX as determined by SPR. Ca2+ has a slight effect on the secondary structure of FIX as determined by circular dichroism spectroscopy. Ca2+ ions are required to maintain in vivo function of FIX Gla domain for its recognition of ACF II. However, Ca2+ at high concentrations (>0.1 mM) inhibits the binding of ACF II to FIX. Ca2+ functions as a switch for the binding between ACF II and FIX. ACF II extends activated partial thromboplastin time more strongly than prothrombin time, suggesting that the binding of ACF II with FIX may play a dominant role in the anticoagulation of ACF II in vivo. © 2012 Wiley Periodicals, Inc. Biopolymers 97: 818–824, 2012.
Co-reporter:Dengke Shen;Xiaolong Xu;Hao Wu;Lili Peng
JBIC Journal of Biological Inorganic Chemistry 2011 Volume 16( Issue 4) pp:523-537
Publication Date(Web):2011 April
DOI:10.1007/s00775-010-0752-9
Anticoagulation factor II (ACF II) isolated from the venom of Agkistrodon acutus is an activated coagulation factor X (FXa)-binding protein with both anticoagulant and hypotensive activities. The thermodynamics of the binding of alkaline earth metal ions to ACF II and their effects on the stability of ACF II and the binding of ACF II to FXa were investigated by isothermal titration calorimetry, fluorescence, differential scanning calorimetry, and surface plasmon resonance. The binding of ACF II to FXa does not have an absolute requirement for Ca2+. Mg2+, Sr2+, and Ba2+ can induce the binding of ACF II to FXa. The radii of the cations bound in ACF II crucially affect the binding affinity of ACF II for cations and the structural stability of ACF II against guanidine hydrochloride and thermal denaturation, whereas the radii of cations bound in FXa markedly affect the binding affinity between ACF II and FXa. The binding affinities of ACF II for cations and the capacities of metal-induced stabilization of ACF II follow the same trend: Ca2+ > Sr2+ > Ba2+. The metal-induced binding affinities of ACF II for FXa follow the trend Mg2+ > Ca2+ > Sr2+ > Ba2+. Although Mg2+ shows significantly low binding affinity with ACF II, Mg2+ is the most effective to induce the binding of ACF II with FXa. Our observations suggest that in blood the bindings of Ca2+ in two sites of ACF II increase the structural stability of ACF II, but these bindings are not essential for the binding of ACF II with FXa, and that the binding of Mg2+ and Ca2+ to FXa may be essential for the recognition between FXa and ACF II. Like Ca2+, the abundant Mg2+ in blood also plays an important role in the anticoagulation of ACF II.
Co-reporter:Hao Wu;Xiaolong Xu;Dengke Shen;Lili Peng
JBIC Journal of Biological Inorganic Chemistry 2011 Volume 16( Issue 1) pp:69-79
Publication Date(Web):2011 January
DOI:10.1007/s00775-010-0703-5
Coagulation factor IX/coagulation factor X binding protein from the venom of Agkistrodon halys Pallas (AHP IX/X-bp) is a unique coagulation factor IX/coagulation factor X binding protein (IX/X-bp). Among all IX/X-bps identified, only AHP IX/X-bp is a Ca2+- and Zn2+-binding protein. The binding properties of Ca2+ and Zn2+ ions binding to apo-AHP IX/X-bp and their effects on the stability of the protein have been investigated by isothermal titration calorimetry, fluorescence spectroscopy, and differential scanning calorimetry. The results show that AHP IX/X-bp has two metal binding sites, one specific for Ca2+ with lower affinity for Zn2+ and one specific for Zn2+ with lower affinity for Ca2+. The bindings of Ca2+ and Zn2+ in the two sites are entropy- and enthalpy-driven. The binding affinity of AHP IX/X-bp for Zn2+ is 1 order of magnitude higher than for Ca2+ for either high-affinity binding or low-affinity binding, which accounts for the existence of one Zn2+ in the purified AHP IX/X-bp. Guanidine hydrochloride (GdnHCl)-induced and thermally induced denaturations of Ca2+–Ca2+-AHP IX/X-bp, Zn2+–Zn2+-AHP IX/X-bp, and Ca2+–Zn2+-AHP IX/X-bp are all a two-state processes with no detectable intermediate state(s), indicating the Ca2+/Zn2+-induced tight packing of the protein. Ca2+ and Zn2+ increase the structural stability of AHP IX/X-bp against GdnHCl or thermal denaturation to a similar extent. Although Ca2+ and Zn2+ have no obvious effect on the secondary structure of AHP IX/X-bp, they induce different rearrangements in local conformation. The Zn2+-stabilized specific conformation of AHP IX/X-bp may be helpful to its recognition of the structure of coagulation factor IX. This work suggests that in vitro, Ca2+ plays a structural rather than an active role in the anticoagulation of AHP IX/X-bp, whereas Zn2+ plays both structural and active roles in the anticoagulation. In blood, Ca2+ binds to AHP IX/X-bp and stabilizes its structure, whereas Zn2+ cannot bind to AHP IX/X-bp owing to the low Zn2+ concentration. AHP IX/X-bp prolongs the clotting time in vivo through its binding only with coagulation factor X/activated coagulation factor X.
Co-reporter:Xiaolong Xu, Liyun Zhang, Zhaofeng Luo, Dengke Shen, Hao Wu, Lili Peng, Jiajia Song and Yan Zhang
Metallomics 2010 vol. 2(Issue 7) pp:480-489
Publication Date(Web):03 Jun 2010
DOI:10.1039/C0MT00017E
AA-NADase from Agkistrodon acutus venom is a unique multicatalytic enzyme with both NADase and AT(D)Pase activities. Among all identified NADases, only AA-NADase contains Cu2+ ions that are essential for its multicatalytic activity. In this study, the interactions between divalent metal ions and AA-NADase and the effects of metal ions on its structure and activity have been investigated by equilibrium dialysis, isothermal titration calorimetry, fluorescence, circular dichroism, dynamic light scattering and HPLC. The results show that AA-NADase has two classes of Cu2+ binding sites, one activator site with high affinity and approximately six inhibitor sites with low affinity. Cu2+ ions function as a switch for its NADase activity. In addition, AA-NADase has one Mn2+ binding site, one Zn2+ binding site, one strong and two weak Co2+ binding sites, and two strong and six weak Ni2+ binding sites. Metal ion binding affinities follow the trend Cu2+ > Ni2+ > Mn2+ > Co2+ > Zn2+, which accounts for the existence of one Cu2+ in the purified AA-NADase. Both NADase and ADPase activities of AA-NADase do not have an absolute requirement for Cu2+, and all tested metal ions activate its NADase and ADPase activities and the activation capacity follows the trend Zn2+ > Mn2+ > Cu2+ ∼ Co2+ > Ni2+. Metal ions serve as regulators for its multicatalytic activity. Although all tested metal ions have no obvious effects on the global structure of AA-NADase, Cu2+- and Zn2+-induced conformational changes around some Trp residues have been observed. Interestingly, each tested metal ion has a very similar activation of both NADase and ADPase activities, suggesting that the two different activities probably occur at the same site.
Co-reporter:Li-Yun Zhang;Zhao-Feng Luo;Hao Wu;Deng-Ke Shen;Li-Li Peng;Yang-Zhong Liu
Biopolymers 2010 Volume 93( Issue 2) pp:141-149
Publication Date(Web):
DOI:10.1002/bip.21312
Abstract
AA-NADase from Agkistrodon acutus venom is a unique multicatalytic enzyme with both NADase and AT(D)Pase activities. Among all identified NADases, only AA-NADase contains Cu(II) and has disulfide-bond linkages between two peptide chains. The effects of the reduction of the disulfide-bonds and Cu(II) in AA-NADase by small-molecule reductants on its NADase and ADPase activities have been investigated by polyacrylamide gel electrophoresis, high performance liquid chromatography, electron paramagnetic resonance spectroscopy and isothermal titration calorimetry. The results show that AA-NADase has six disulfide-bonds and fifteen free cysteine residues. L-ascorbate inhibits AA-NADase on both NADase and ADPase activities through the reduction of Cu(II) in AA-NADase to Cu(I), while other reductants, dithiothreitol, glutathione and tris(2-carboxyethyl)phosphine inhibit both NADase and ADPase activities through the reduction of Cu(II) to Cu(I) and the cleavage of disulfide-bonds in AA-NADase. Apo-AA-NADase can recover its NADase and ADPase activities in the presence of 1 mM Zn(II). However, apo-AA-NADase does not recover any NADase or ADPase activity in the presence of 1 mM Zn(II) and 2 mM TCEP. The multicatalytic activity relies on both disulfide-bonds and Cu(II), while Cu(I) can not activate the enzyme activities. AA-NADase is probably only active as a dimer. The inhibition curves for both ADPase and NADase activities by each reductant share a similar trend, suggesting both ADPase and NADase activities probably occur at the same site. In addition, we also find that glutathione and L-ascorbate are endogenous inhibitors to the multicatalytic activity of AA-NADase. © 2009 Wiley Periodicals, Inc. Biopolymers 93: 141–149, 2010.
This article was originally published online as an accepted preprint. The “Published Online” date corresponds to the preprint version. You can request a copy of the preprint by emailing the Biopolymers editorial office at biopolymers@wiley.com
Co-reporter:Xiaolong Xu;Liyun Zhang;Dengke Shen
JBIC Journal of Biological Inorganic Chemistry 2009 Volume 14( Issue 4) pp:559-571
Publication Date(Web):2009 May
DOI:10.1007/s00775-009-0470-3
Anticoagulation factor I (ACF I) isolated from the venom of Agkistrodon acutus is an activated coagulation factor X (FXa)-binding protein that binds in a Ca2+-dependent fashion with marked anticoagulant activity. The thermodynamics of the binding of alkaline earth metal ions to ACF I and the effects of alkaline earth metal ions on the guanidine hydrochloride (GdnHCl)-induced unfolding of ACF I and the binding of ACF I to FXa were studied by isothermal titration calorimetry, fluorescence, circular dichroism, and surface plasmon resonance, respectively. The results indicate that the ionic radii of the cations occupying Ca2+-binding sites in ACF I crucially affect the binding affinity of ACF I for alkaline earth metal ions as well as the structural stability of ACF I against GdnHCl denaturation. Sr2+ and Ba2+, with ionic radii larger than the ionic radius of Ca2+, can bind to Ca2+-free ACF I (apo-ACF I), while Mg2+, with an ionic radius smaller than that of Ca2+, shows significantly low affinity for the binding to apo-ACF I. All bindings of Ca2+, Sr2+, and Ba2+ ions in two sites of ACF I are mainly enthalpy-driven and the entropy is unfavorable for them. Sr2+-stabilized ACF I exhibits slightly lower resistance to GdnHCl denaturation than Ca2+–ACF I, while Ba2+-stabilized ACF I exhibits much lower resistance to GdnHCl denaturation than Ca2+–ACF I. Mg2+ and Sr2+, with ionic radii close to that of Ca2+, can bind to FXa and therefore also induce the binding of ACF I to FXa, whereas Ba2+, with a much larger ionic radius than Ca2+, cannot support the binding of ACF I with FXa. Our observations suggest that bindings of Ca2+, Sr2+, and Ba2+ ions in two sites of ACF I increase the structural stability of ACF I, but these bindings are not essential for the binding of ACF I with FXa, and that the binding of Mg2+, Ca2+, and Sr2+ ions to FXa may be essential for the recognition between FXa and ACF I.
Co-reporter:Xiaolong Xu;Jiexia Chen;Liyun Zhang;Shouye Wang;Dengke Shen
Journal of Fluorescence 2007 Volume 17( Issue 2) pp:215-221
Publication Date(Web):2007 March
DOI:10.1007/s10895-007-0160-x
Agkisacutacin isolated from the venom of Agkistrodon acutus is a coagulation factor IX / coagulation factor X-binding protein with marked anticoagulant- and platelet-modulating activities. Ca2+ ion-induced stabilization and refolding of Agkisacutacin have been studied by following fluorescent measurements. Ca2+ ions not only increase the structural stability of agkisacutacin against GdnHCl denaturation, but also induce its refolding. The GdnHCl-induced unfolding of the apo-agkisacutacin and the purified agkisacutacin is a single-step process with no detectable intermediate state. Ca2+ ions play an important role in the stabilization of the structure of agkisacutacin. Ca2+-stabilized agkisacutacin exhibits higher resistance to GdnHCl denaturation than the apo-agkisacutacin. It is possible to induce refolding of the unfolded apo-agkisacutacin merely by adding 1 mM Ca2+ ions without changing the concentration of the denaturant. The kinetic result of Ca2+-induced refolding provides evidences for that agkisacutacin consists of at least two refolding phases and the first phase of Ca2+-induced refolding should involve the formation of the compact Ca2+-binding site regions, and subsequently, the protein undergoes further conformational rearrangements to form the native structure.
Co-reporter:Xiaolong Xu;Jiexia Chen;Liyun Zhang;Qingliang Liu
Biopolymers 2007 Volume 85(Issue 1) pp:
Publication Date(Web):24 OCT 2006
DOI:10.1002/bip.20617
Acutolysin A isolated from the venom of Agkistrodon acutus is a protein of 22 kDa with marked haemorrhagic and proteolytic activities. The metal ions- and pH-induced conformational changes of acutolysin A have been studied by following fluorescence and activity measurements. Here, we provide evidence for the fact that native holo-acutolysin A adopts two subtly different conformations, native state a (Na) stable in the weak acidic pH range from 6.0 to 7.0 with low activity and native state b (Nb) stable in the weak alkaline pH range from 7.5 to 9.0 with high activity. Holo-acutolysin A has an optimum pH of 8.5 for caseinolytic activity, and the protein adopts the most stable conformation with the maximum fluorescence at pH 8.5. The Ca2+ and Zn2+ ions have significant effects on both the pH-induced denaturing transition curve and the pH-dependent activity curve. Addition of 1 mM Ca2+ to holo-acutolysin A shifts both the acid-induced denaturing transition curve and the end zone of acid-induced inactivation curve towards lower pH value, and shifts both the alkali-induced denaturing transition curve and the end zone of alkali-induced inactivation curve towards higher pH value. Addition of 1 mM Zn2+ also shifts both the alkali-induced denaturing transition curve and the end zone of alkali-induced inactivation curve towards higher pH value and shifts the acid-induced denaturing transition curve to lower pH value, but has little effect on the acid-induced inactivation. Removal of Ca2+ and Zn2+ from the protein enhances its sensitivity to pH and significantly reduces its overall stability during acid-induced denaturation. It is also evident from the present work that the free Zn2+-induced inactivation in the pH range from 8.0 to 9.0 should be attributed to the effect of Zn(OH)2 precipitation on the protein. © 2006 Wiley Periodicals, Inc. Biopolymers 85: 81–90, 2007.
This article was originally published online as an accepted preprint. The “Published Online” date corresponds to the preprint version. You can request a copy of the preprint by emailing the Biopolymers editorial office at biopolymers@wiley.com
Co-reporter:Xiao-Long Xu;Jie-Xia Chen;Li-Yun Zhang;Xiang-Hu Liu;Wen-Qi Liu;Qing-Liang Liu
Biopolymers 2006 Volume 82(Issue 2) pp:
Publication Date(Web):10 FEB 2006
DOI:10.1002/bip.20476
Anticoagulation factor II (ACF II) isolated from the venom of Agkistrodon acutus is an activated coagulation factor X-binding protein in a Ca2+-dependent fashion with marked anticoagulant activity. The equilibrium unfolding of rare earth ions (RE3+)-reconstituted ACF II in guanidine hydrochloride (GdnHCl) solution was studied by fluorescence. The GdnHCl-induced unfolding of RE3+ (Nd3+, Sm3+, Eu3+, Gd3+)-reconstituted ACF II follows a three-state transition with a stable intermediate state. Substitutions of the RE3+ ions for Ca2+ in ACF II decrease the conformational stability of its native state but markedly increase the conformational stability of its intermediate state. The free energy change of RE3+–ACF II from the intermediate state to denatured state linearly increases with the increase of ionic potentials of bound metal ions (Ca2+, Nd3+, Sm3+, Eu3+, and Gd3+). The refolding of ACF II from the unfolded state to the intermediate state can be induced merely by adding 10 μM RE3+ ions without changing the concentration of the denaturant. The kinetic results of the RE3+-induced refolding provide evidence indicating that the intermediate state of RE3+–ACF II consists of at least two refolding phases and that the refolding rate constant values of the faster phase decrease with the increase of the difference between the radii of Ca2+ and RE3+, but the refolding rate constant values of the slower phase are similar to each other. The results of this study indicate that the size of metal ion is the major factor responsible for the metal ion-induced conformational stabilization of the native ACF II, while the metal ionic potential plays a predominant role in stabilizing the conformation for the intermediate state. © 2006 Wiley Periodicals, Inc. Biopolymers 82: 167–175, 2006
This article was originally published online as an accepted preprint. The “Published Online” date corresponds to the preprint version. You can request a copy of the preprint by emailing the Biopolymers editorial office at biopolymers@wiley.com
Co-reporter:Shasha Wang, Xiaolong Xu, Shang Gao, Shanshan Zhu, Rui Rong, Bing Li
Protein Expression and Purification (July 2014) Volume 99() pp:99-105
Publication Date(Web):1 July 2014
DOI:10.1016/j.pep.2014.04.007
•A non-hemorrhagic fibrinogenase (DAnase) was purified from Deinagkistrodon acutus venom.•DAnase was a disulfide-linked dimer requiring disulfide bridge(s) for its activity.•DAnase had arginine esterase activity and inhibited ADP-induced platelet aggregation.•DAnase was devoid of hemorrhagic activity and Factor XIII activation activity.•DAnase has a potential clinical application for the therapy of thrombosis disease.A novel fibrinogenase, DAnase, was purified from the venom of Deinagkistrodon acutus by a combination of anion and cation exchange chromatography. Unlike other fibrinogenases which are usually single polypeptide chain proteins, the enzyme was a disulfide-linked dimer with an isoelectric point of 6.03 and an apparent molecular weight of 25 kDa on SDS–polyacrylamide gel electrophoresis. DAnase showed α-fibrinogenase activity devoid of fibrinolytic activity. It hydrolyzed rapidly the Aα-chain of fibrinogen and followed by the Bβ-chain and did not cleave the γ-chain. It also exhibited arginine esterase activity. The fibrinogenolytic and arginine esterase activities were completely inhibited by phenylmethanesulfonyl fluoride or tris-(2-carboxyethyl)phosphine hydrochloride, but not by EDTA, indicating that DAnase is a serine protease requiring disulfide bridge(s) for its activity. The protease strongly inhibited ADP-induced platelet aggregation in human platelet-rich plasma but was lack of ADPase activity, indicating that its fibrinogenolytic activity is involved in its inhibition of ADP-induced platelet aggregation. DAnase was devoid of hemorrhagic activity and Factor XIII activation activity. DAnase may have a potential clinical application for the therapy of thrombosis disease.
Co-reporter:Hao Wu, Xiaolong Xu, Liyun Zhang, Dengke Shen, Lili Peng, Yan Zhang, Jiajia Song
Toxicon (15 June 2010) Volume 55(Issue 7) pp:1358-1364
Publication Date(Web):15 June 2010
DOI:10.1016/j.toxicon.2010.02.010
Factor IX-binding protein (AHP IX-bp), a Ca2+- and Zn2+-binding protein from the venom of Agkistrodon Halys Pallas was reported to bind specifically with factor IX in a Zn2+-dependent manner. Here we have purified AHP IX-bp by a simple two-step of chromatography procedure and found that AHP IX-bp also binds factor Xa (FXa) with high binding-affinity in a Mg2+-dependent manner. Although Mg2+ ions have a significantly low binding-affinity for apo-AHP IX-bp as determined by isothermal titration calorimetry, they can induce the binding of apo-AHP IX-bp with FXa even in the absence of Ca2+ as determined by native PAGE and surface plasmon resonance. Mg2+ ions are required to maintain in vivo function of FX Gla domain for its recognition of AHP IX-bp. Both Ca2+ and Zn2+ ions fail to induce the binding between apo-AHP IX-bp and FXa. The abundant Mg2+ ions in blood play an important role in the anticoagulation of AHP IX-bp.
Co-reporter:Yan Zhang, Xiaolong Xu, Dengke Shen, Jiajia Song, Mingchun Guo, Xincheng Yan
Toxicon (1 June 2012) Volume 59(Issues 7–8) pp:718-723
Publication Date(Web):1 June 2012
DOI:10.1016/j.toxicon.2012.03.006
Anticoagulation factor I (ACF I), a snake C-type lectin (snaclec) from the venom of Agkistrodon acutus binds specifically with activated factor X (FXa) in a Ca2+-dependent manner and prolongs the blood-clotting time in vitro. In this study, the inhibition of the coagulation pathway by ACF I was measured in vivo by activated partial thromboplastin time and prothrombin time assays and the binding of ACF I to factor IX (FIX) was investigated by native PAGE and surface plasmon resonance. The results indicate that ACF I inhibits both intrinsic and extrinsic coagulation pathways, but does not inhibit thrombin activity. ACF I also binds FIX in a Ca2+-dependent manner and their maximal binding occurs at 0.25 mM Ca2+. ACF I has a higher binding-affinity to FIX than to FX. Ca2+ is required to maintain in vivo function of FIX Gla domain for its recognition of ACF I. However, Ca2+ at high concentrations (>0.25 mM) inhibits the binding of ACF I to FIX. Ca2+ functions as a switch for the binding between ACF I and FIX. The results suggest that the binding of ACF I with FIX may play a dominant role in the anticoagulation activity of ACF I in vivo.Highlights► ACF I inhibits both intrinsic and extrinsic coagulation pathways, but does not inhibit thrombin activity. ► ACF I binds FIX in a Ca2+-dependent manner and the maximal binding between ACF I and FIX occurs at 0.25 mM Ca2+. ► Ca2+ functions as a switch for the binding between ACF I and FIX. ► ACF I has been identified as a FIX and FX binding protein with a higher binding-affinity to FIX than to FX. ► The binding of ACF I with FIX plays a dominant role in the anticoagulation of ACF I in vivo.