Co-reporter:Ashwani Kumar Vashishtha and Robert D. Kuchta
Biochemistry 2016 Volume 55(Issue 7) pp:1168-1177
Publication Date(Web):February 2, 2016
DOI:10.1021/acs.biochem.6b00065
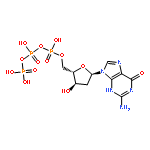
We examined the impact of two clinically approved anti-herpes drugs, acyclovir and Forscarnet (phosphonoformate), on the exonuclease activity of the herpes simplex virus-1 DNA polymerase, UL30. Acyclovir triphosphate and Foscarnet, along with the closely related phosphonoacetic acid, did not affect exonuclease activity on single-stranded DNA. Furthermore, blocking the polymerase active site due to either binding of Foscarnet or phosphonoacetic acid to the E–DNA complex or polymerization of acyclovir onto the DNA also had a minimal effect on exonuclease activity. The inability of the exonuclease to excise acyclovir from the primer 3′-terminus results from the altered sugar structure directly impeding phosphodiester bond hydrolysis as opposed to inhibiting binding, unwinding of the DNA by the exonuclease, or transfer of the DNA from the polymerase to the exonuclease. Removing the 3′-hydroxyl or the 2′-carbon from the nucleotide at the 3′-terminus of the primer strongly inhibited exonuclease activity, although addition of a 2′-hydroxyl did not affect exonuclease activity. The biological consequences of these results are twofold. First, the ability of acyclovir and Foscarnet to block dNTP polymerization without impacting exonuclease activity raises the possibility that their effects on herpes replication may involve both direct inhibition of dNTP polymerization and exonuclease-mediated destruction of herpes DNA. Second, the ability of the exonuclease to rapidly remove a ribonucleotide at the primer 3′-terminus in combination with the polymerase not efficiently adding dNTPs onto this primer provides a novel mechanism by which the herpes replication machinery can prevent incorporation of ribonucleotides into newly synthesized DNA.
Co-reporter:Ashwani Kumar Vashishtha and Robert D. Kuchta
Biochemistry 2015 Volume 54(Issue 2) pp:240-249
Publication Date(Web):December 17, 2014
DOI:10.1021/bi500840v
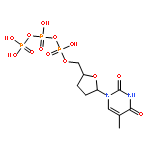
The herpes polymerase–processivity factor complex consists of the catalytic UL30 subunit containing both polymerase and proofreading exonuclease activities and the UL42 subunit that acts as a processivity factor. Curiously, the highly active exonuclease has minimal impact on the accumulation of mismatches generated by the polymerase activity. We utilized a series of oligonucleotides of defined sequence to define the interactions between the polymerase and exonuclease active sites. Exonuclease activity requires unwinding of two nucleotides of the duplex primer–template. Surprisingly, even though the exonuclease rate is much higher than the rate of DNA dissociation, the exonuclease degrades both single- and double-stranded DNA in a nonprocessive manner. Efficient proofreading of incorrect nucleotides incorporated by the polymerase would seem to require efficient translocation of DNA between the exonuclease and polymerase active sites. However, we found that translocation of DNA from the exonuclease to polymerase active site is remarkably inefficient. Consistent with inefficient translocation, the DNA binding sites for the exonuclease and polymerase active sites appear to be largely independent, such that the two activities appear noncoordinated. Finally, the presence or absence of UL42 did not impact the coordination of the polymerase and exonuclease activities. In addition to providing fundamental insights into how the polymerase and exonuclease function together, these activities provide a rationale for understanding why the exonuclease minimally impacts accumulation of mismatches by the purified polymerase and raise the question of how these two activities function together in vivo.
Co-reporter:Andrew C. Olson ; Jennifer N. Patro ; Milan Urban
Journal of the American Chemical Society 2013 Volume 135(Issue 4) pp:1205-1208
Publication Date(Web):January 14, 2013
DOI:10.1021/ja309866m
To better understand the energetics of accurate DNA replication, we directly measured ΔGo for the incorporation of a nucleotide into elongating dsDNA in solution (ΔGoincorporation). Direct measurements of the energetic difference between synthesis of correct and incorrect base pairs found it to be much larger than previously believed (average ΔΔGoincorporation = 5.2 ± 1.34 kcal mol–1). Importantly, these direct measurements indicate that ΔΔGoincorporation alone can account for the energy required for highly accurate DNA replication. Evolutionarily, these results indicate that the earliest polymerases did not have to evolve sophisticated mechanisms to replicate nucleic acids; they may only have had to take advantage of the inherently more favorable ΔGo for polymerization of correct nucleotides. These results also provide a basis for understanding how polymerases replicate DNA (or RNA) with high fidelity.
Co-reporter:Travis J. Lund, Nisha A. Cavanaugh, Nicolas Joubert, Milan Urban, Jennifer N. Patro, Michal Hocek, and Robert D. Kuchta
Biochemistry 2011 Volume 50(Issue 33) pp:
Publication Date(Web):July 15, 2011
DOI:10.1021/bi2006916
We utilized a series of pyrimidine analogues modified at O2, N-3, and N4/O4 to determine if two B family DNA polymerases, human DNA polymerase α and herpes simplex virus I DNA polymerase, choose whether to polymerize pyrimidine dNTPs using the same mechanisms they use for purine dNTPs. Removing O2 of a pyrimidine dNTP vastly decreased the level of incorporation by these enzymes and also compromised fidelity in the case of C analogues, while removing O2 from the templating base had more modest effects. Removing the Watson–Crick hydrogen bonding groups of N-3 and N4/O4 greatly impaired polymerization, both of the resulting dNTP analogues and of natural dNTPs opposite these pyrimidine analogues when present in the template strand. Thus, the Watson–Crick hydrogen bonding groups of a pyrimidine clearly play an important role in enhancing correct dNTP polymerization but are not essential for preventing misincorporation. These studies also indicate that DNA polymerases recognize bases extremely asymmetrically, both in terms of whether they are a purine or pyrimidine and whether they are in the template or are the incoming dNTP. The mechanistic implications of these results with regard to how polymerases discriminate between right and wrong dNTPs are discussed.
Co-reporter:Gudrun Stengel, Byron W. Purse, Robert D. Kuchta
Analytical Biochemistry 2011 Volume 416(Issue 1) pp:53-60
Publication Date(Web):1 September 2011
DOI:10.1016/j.ab.2011.04.033
The cytosine analogs 1,3-diaza-2-oxophenothiazine (tC) and 1,3-diaza-2-oxophenoxazine (tCo) stand out among fluorescent bases due to their unquenched fluorescence emission in double-stranded DNA. Recently, we reported a method for the generation of densely tCo-labeled DNA by polymerase chain reaction (PCR) that relied on the use of the extremely thermostable Deep Vent polymerase. We have now developed a protocol that employs the more commonly used Taq polymerase. Supplementing the PCR with Mn2+ or Co2+ ions dramatically increased the amount of tCo triphosphate (dtCoTP) incorporated and, thus, enhanced the brightness of the PCR products. The resulting PCR products could be easily detected in gels based on their intrinsic fluorescence. The Mn2+ ions modulate the PCR by improving the bypass of template tCo and the overall catalytic efficiency. In contrast to the lower fidelity during tCo bypass, Mn2+ improved the ability of Taq polymerase to distinguish between dtCoTP and dTTP when copying a template dA. Interestingly, Mn2+ ions hardly affect the fluorescence emission of tC(o), whereas the coordination of Co2+ ions with the phosphate groups of DNA and nucleotides statically quenches tC(o) fluorescence with small reciprocal Stern–Vollmer constants of 10–300 μM.
Co-reporter:Gudrun Stengel, Milan Urban, Byron W. Purse and Robert D. Kuchta
Analytical Chemistry 2010 Volume 82(Issue 3) pp:1082
Publication Date(Web):January 12, 2010
DOI:10.1021/ac902456n
Fluorescent RNA is an important analytical tool in medical diagnostics, RNA cytochemistry, and RNA aptamer development. We have synthesized the fluorescent ribonucleotide analogue 1,3-diaza-2-oxophenothiazine-ribose-5′-triphosphate (tCTP) and tested it as substrate for T7 RNA polymerase in transcription reactions, a convenient route for generating RNA in vitro. When transcribing a guanine, T7 RNA polymerase incorporates tCTP with 2-fold higher catalytic efficiency than CTP and efficiently polymerizes additional NTPs onto the tC. Remarkably, T7 RNA polymerase does not incorporate tCTP with the same ambivalence opposite guanine and adenine with which DNA polymerases incorporate the analogous dtCTP. While several DNA polymerases discriminated against a d(tC-A) base pair only by factors <10, T7 RNA polymerase discriminates against tC-A base pair formation by factors of 40 and 300 when operating in the elongation and initiation mode, respectively. These catalytic properties make T7 RNA polymerase an ideal tool for synthesizing large fluorescent RNA, as we demonstrated by generating a ∼800 nucleotide RNA in which every cytosine was replaced with tC.
Co-reporter:Andrew C. Olson, Eric Rosenblum, and Robert D. Kuchta
Biochemistry 2010 Volume 49(Issue 47) pp:
Publication Date(Web):October 29, 2010
DOI:10.1021/bi101011j
The influenza RNA-dependent RNA polymerase (RdRp) both replicates the flu’s RNA genome and transcribes its mRNA. Replication occurs de novo; however, initiation of transcription requires a 7-methylguanosine 5′-capped primer that is “snatched” from host mRNA via endonuclease and cap binding functions of the influenza polymerase. A key question is how the virus regulates the relative amounts of transcription and replication. We found that the concentration of a capped cellular mRNA, the concentration of the 5′ end of the viral RNA, and the concentration of RdRp all regulate the relative amounts of replication versus transcription. The host mRNA, from which the RdRp snatches its capped primer, acts to upregulate transcription and repress replication. Elevated concentrations of the RdRp itself switch the influenza polymerase toward replication, likely through an oligomerization of the polymerase. The 5′ end of the vRNA template both activates replication and inhibits transcription of the vRNA template, thereby indicating that RdRp contains an allosteric binding site for the 5′ end of the vRNA template. These data provide insights into the regulation of RdRp throughout the viral life cycle and how it synthesizes the appropriate amounts of viral mRNA and replication products (vRNA and cRNA).
Co-reporter:Milan Urban, Nicolas Joubert, Byron W. Purse, Michal Hocek and Robert D. Kuchta
Biochemistry 2010 Volume 49(Issue 4) pp:
Publication Date(Web):December 23, 2009
DOI:10.1021/bi9019516
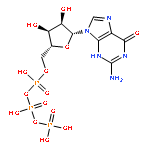
Human DNA primase synthesizes short RNA primers that DNA polymerase α then elongates during the initiation of all new DNA strands. Even though primase misincorporates NTPs at a relatively high frequency, this likely does not impact the final DNA product since the RNA primer is replaced with DNA. We used an extensive series of purine and pyrimidine analogues to provide further insights into the mechanism by which primase chooses whether or not to polymerize a NTP. Primase readily polymerized a size-expanded cytosine analogue, 1,3-diaza-2-oxophenothiazine NTP, across from a templating G but not across from A. The enzyme did not efficiently polymerize NTPs incapable of forming two Watson−Crick hydrogen bonds with the templating base with the exception of UTP opposite purine deoxyribonucleoside. Likewise, primase did not generate base pairs between two nucleotides with altered Watson−Crick hydrogen-bonding patterns. Examining the mechanism of NTP polymerization revealed that human primase can misincorporate NTPs via both template misreading and a primer-template slippage mechanism. Together, these data demonstrate that human primase strongly depends on Watson−Crick hydrogen bonds for efficient nucleotide polymerization, much more so than the mechanistically related herpes primase, and provide insights into the potential roles of primer-template stability and base tautomerization during misincorporation.
Co-reporter:Gudrun Stengel, Milan Urban, Byron W. Purse and Robert D. Kuchta
Analytical Chemistry 2009 Volume 81(Issue 21) pp:9079
Publication Date(Web):October 7, 2009
DOI:10.1021/ac9017555
Fluorescent DNA of high molecular weight is an important tool for studying the physical properties of DNA and DNA−protein interactions, and it plays a key role in modern biotechnology for DNA sequencing and detection. While several DNA polymerases can incorporate large numbers of dye-linked nucleotides into primed DNA templates, the amplification of the resulting densely labeled DNA strands by polymerase chain reaction (PCR) is problematic. Here, we report a method for high density labeling of DNA in PCR reactions employing the 5′-triphosphate of 1,3-diaza-2-oxo-phenoxazine (tCo) and Deep Vent DNA polymerase. tCo is a fluorescent cytosine analogue that absorbs and emits light at 365 and 460 nm, respectively. We obtained PCR products that were fluorescent enough to directly visualize them in a gel by excitation with long UV light, thus eliminating the need for staining with ethidium bromide. Reactions with Taq polymerase failed to produce PCR products in the presence of only small amounts of dtCoTP. A comparative kinetic study of Taq and Deep Vent polymerase revealed that Taq polymerase, although it inserts dtCoTP with high efficiency opposite G, is prone to forming mutagenic tCo-A base pairs and does not efficiently extend base pairs containing tCo. These kinetics features explain the poor outcome of the PCR reactions with Taq polymerase. Since tCo substitutes structurally for cytosine, the presented labeling method is believed to be less invasive than labeling with dye-linked nucleotides and, therefore, produces DNA that is ideally suited for biophysical studies.
Co-reporter:Jennifer N. Patro, Milan Urban and Robert D. Kuchta
Biochemistry 2009 Volume 48(Issue 1) pp:
Publication Date(Web):December 12, 2008
DOI:10.1021/bi801823z
We used a series of dNTP analogues in conjunction with templates containing modified bases to elucidate the role that N2 of a purine plays during dNTP polymerization by human DNA polymerase α. Removing N2 from dGTP had small effects during correct incorporation opposite C but specifically increased misincorporation opposite A. Adding N2 to dATP and related analogues had small and variable effects on the efficiency of polymerization opposite T. However, the presence of N2 greatly enhanced polymerization of these dATP analogues opposite a template C. The ability of N2 to enhance polymerization opposite C likely results from formation of a hydrogen bond between the purine N2 and pyrimidine O2. Even in those cases where formation of a wobble base pair, tautomerization, and/or protonation of the base pair between the incoming dNTP and template base cannot occur (e.g., 2-pyridone·purine (or purine analogue) base pairs), N2 enhanced formation of the base pair. Importantly, N2 had similar effects on dNTP polymerization both when added to the incoming purine dNTP and when added to the template base being replicated. The mechanistic implications of these results regarding how pol α discriminates between right and wrong dNTPs are discussed.
Co-reporter:Nisha A. Cavanaugh, Milan Urban, Jeffrey Beckman, Thomas E. Spratt and Robert D. Kuchta
Biochemistry 2009 Volume 48(Issue 15) pp:
Publication Date(Web):January 23, 2009
DOI:10.1021/bi8022202
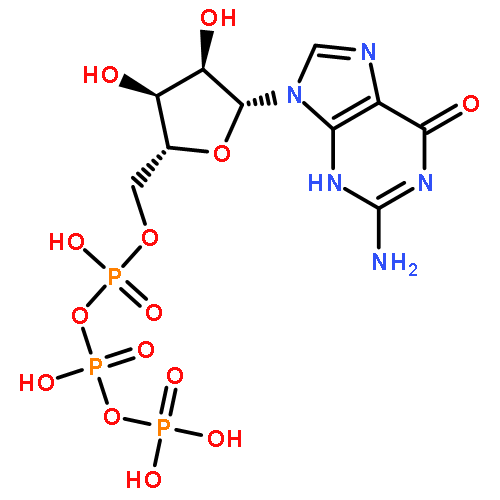
To accurately replicate its viral genome, the Herpes Simplex Virus 1 (HSV-1) DNA polymerase usually polymerizes the correct natural 2′-deoxy-5′-triphosphate (dNTP) opposite the template base being replicated. We employed a series of purine−dNTP analogues to determine the chemical features of the base necessary for the herpes polymerase to avoid polymerizing incorrect dNTPs. The enzyme uses N-3 to prevent misincorporation of purine dNTPs but does not require N-3 for correct polymerization. A free pair of electrons on N-1 also helps prevent misincorporation opposite A, C, and G and strongly drives polymerization opposite T. N6 contributes a small amount both for preventing misincorporation and for correct polymerization. Within the context of guanine in either the incoming dNTP or the template base being replicated, N2 prevents misincorporation opposite adenine but plays at most a minor role for incorporation opposite C. In contrast, adding N2 to the dNTPs of either adenine, purine, 6-chloropurine, or 1-deazapurine greatly enhances incorporation opposite C, likely via the formation of a hydrogen bond between N2 of the purine and O2 of the pyrimidine. Herpes polymerase is very sensitive to the structure of the base pair at the primer 3′-terminus since eliminating N-1, N-3, or N6 from a purine nucleotide at the primer 3′-terminus interfered with polymerization of the next two dNTPs. The biological and evolutionary implications of these data are discussed.
Co-reporter:Michael Trostler, Alison Delier, Jeff Beckman, Milan Urban, Jennifer N. Patro, Thomas E. Spratt, Lorena S. Beese and Robert D. Kuchta
Biochemistry 2009 Volume 48(Issue 21) pp:
Publication Date(Web):April 6, 2009
DOI:10.1021/bi900104n
We used a series of dATP and dGTP analogues to determine how DNA polymerase I from Bacillus stearothermophilus (BF), a prototypical A family polymerase, uses N-1, N2, N-3, and N6 of purine dNTPs to differentiate between right and wrong nucleotide incorporation. Altering any of these nitrogens had two effects. First, it decreased the efficiency of correct incorporation of the resulting dNTP analogue, with the loss of N-1 and N-3 having the most severe effects. Second, it dramatically increased the rate of misincorporation of the resulting dNTP analogues, with alterations in either N-1 or N6 having the most severe impacts. Adding N2 to dNTPs containing the bases adenine and purine increased the degree of polymerization opposite T but also tremendously increased the degree of misincorporation opposite A, C, and G. Thus, BF uses N-1, N2, N-3, and N6 of purine dNTPs both as negative selectors to prevent misincorporation and as positive selectors to enhance correct incorporation. Comparing how BF discriminates between right and wrong dNTPs with both B family polymerases and low-fidelity polymerases indicates that BF has chosen a unique solution vis-à-vis these other enzymes and, therefore, that nature has evolved at least three mechanistically distinct solutions.
Co-reporter:Gudrun Stengel, Byron W. Purse, L. Marcus Wilhelmsson, Milan Urban and Robert D. Kuchta
Biochemistry 2009 Volume 48(Issue 31) pp:
Publication Date(Web):July 6, 2009
DOI:10.1021/bi9006995
We studied the incorporation of the fluorescent cytidine analogues 1,3-diaza-2-oxophenothiazine (tC) and 1,3-diaza-2-oxophenoxazine (tCo) by human DNA polymerase α and Klenow fragment of DNA polymerase I (Escherichia coli). These tricyclic nucleobases possess the regular hydrogen bonding interface of cytosine but are significantly expanded in size toward the major groove. Despite the size alteration, both DNA polymerases insert dtCTP and dtCoTP with remarkable catalytic efficiency. Polymerization opposite guanine is comparable to the insertion of dCTP, while the insertion opposite adenine is only ∼4−11 times less efficient than the formation of a T-A base pair. Both enzymes readily extend the formed tC(o)-G and tC(o)-A base pairs and can incorporate at least four consecutive nucleotide analogues. Consistent with these results, both DNA polymerases efficiently polymerize dGTP and dATP when tC and tCo are in the template strand. Klenow fragment inserts dGTP with a 4−9-fold higher probability than dATP, while polymerase α favors dGTP over dATP by a factor of 30−65. Overall, the properties of tC(o) as a templating base and as an incoming nucleotide are surprisingly symmetrical and may be universal for A and B family DNA polymerases. This finding suggests that the aptitude for ambivalent base pairing is a consequence of the electronic properties of tC(o).
Co-reporter:Jennifer N. Patro, Milan Urban and Robert D. Kuchta
Biochemistry 2009 Volume 48(Issue 34) pp:
Publication Date(Web):July 30, 2009
DOI:10.1021/bi900777s
To better understand how DNA polymerases interact with mutagenic bases, we examined how human DNA polymerase α (pol α), a B family enzyme, and DNA polymerase from Bacillus stearothermophilus (BF), an A family enzyme, generate adenine:hypoxanthine and adenine:8-oxo-7,8-dihydroguanine (8-oxoG) base pairs. Pol α strongly discriminated against polymerizing dATP opposite 8-oxoG, and removing N1, N6, or N7 further inhibited incorporation, whereas removing N3 from dATP dramatically increased incorporation (32-fold). Eliminating N6 from 3-deaza-dATP now greatly reduced incorporation, suggesting that incorporation of dATP (analogues) opposite 8-oxoguanine proceeds via a Hoogsteen base pair and that pol α uses N3 of a purine dNTP to block this incorporation. Pol α also polymerized 8-oxo-dGTP across from a templating A, and removing N6 from the template adenine inhibited incorporation of 8-oxoG. The effects of N1, N6, and N7 demonstrated a strong interdependence during formation of adenine:hypoxanthine base pairs by pol α, and N3 of dATP again helps prevent polymerization opposite a templating hypoxanthine. BF very efficiently polymerized 8-oxo-dGTP opposite adenine, and N1 and N7 of adenine appear to play important roles. BF incorporates dATP opposite 8-oxoG less efficiently, and modifying N1, N6, or N7 greatly inhibits incorporation. N6 and, to a lesser extent, N1 help drive hypoxanthine:adenine base-pair formation by BF. The mechanistic implications of these results showing that different polymerases interact very differently with base lesions are discussed.
Co-reporter:Nisha A. Cavanaugh, Kathryn A. Ramirez-Aguilar, Milan Urban and Robert D. Kuchta
Biochemistry 2009 Volume 48(Issue 43) pp:
Publication Date(Web):September 30, 2009
DOI:10.1021/bi9010144
The helicase−primase complex from herpes simplex virus-1 contains three subunits, UL5, UL52, and UL8. We generated each of the potential two-subunit complexes, UL5−UL52, UL5−UL8, and UL52−UL8, and used a series of kinetic and photo-cross-linking studies to provide further insights into the roles of each subunit in DNA binding and primer synthesis. UL8 increases the rate of primer synthesis by UL5−UL52 by increasing the rate of primer initiation (two NTPs → pppNpN), the rate-limiting step in primer synthesis. The UL5−UL8 complex lacked any detectable catalytic activity (DNA-dependent ATPase, primase, or RNA polymerase using a RNA primer−template and NTPs as substrates) but could still bind DNA, indicating that UL52 must provide some key amino acids needed for helicase function. The UL52−UL8 complex lacked detectable DNA-dependent ATPase activity and could not synthesize primers on single-stranded DNA. However, it exhibited robust RNA polymerase activity using a RNA primer−template and NTPs as substrates. Thus, UL52 must contain the entire primase active site needed for phosphodiester bond formation, while UL5 minimally contributes amino acids needed for the initiation of primer synthesis. Photo-cross-linking experiments using single-stranded templates containing 5-iodouracil either before, in, or after the canonical 3′-GPyPy (Py is T or C) initiation site for primer synthesis showed that only UL5 cross-linked to the DNA. This occurred for the UL5−UL52, UL5−UL52−UL8, and UL5−UL8 complexes and whether the reaction mixtures contained NTPs. Photo-cross-linking of a RNA primer−template, the product of primer synthesis, containing 5-iodouracil in the template generated the same apparent cross-linked species.
Co-reporter:Milan Urban, Nicolas Joubert, Michal Hocek, Richard E. Alexander and Robert D. Kuchta
Biochemistry 2009 Volume 48(Issue 46) pp:
Publication Date(Web):October 17, 2009
DOI:10.1021/bi901476k
Herpes simplex virus-1 primase misincorporates the natural NTPs at frequencies of around one error per 30 NTPs polymerized, making it one of the least accurate polymerases known. We used a series of nucleotide analogues to further test the hypothesis that primase requires Watson−Crick hydrogen bond formation to efficiently polymerize a NTP. Primase could not generate base pairs containing a complete set of hydrogen bonds in an altered arrangement (isoguanine·isocytosine) and did not efficiently polymerize dNTPs completely incapable of forming Watson−Crick hydrogen bonds opposite templating bases incapable of forming Watson−Crick hydrogen bonds. Similarly, primase did not incorporate most NTPs containing hydrophobic bases incapable of Watson−Crick hydrogen bonding opposite natural template bases. However, 2-pyridone NTP and 4-methyl-2-pyridone NTP provided striking exceptions to this rule. The effects of removing single Watson−Crick hydrogen bonding groups from either the NTP or templating bases varied from almost no effect to completely blocking polymerization depending both on the parental base pair (G·C vs A·T/U) and which base pair of the growing primer (second, third, or fourth) was examined. Thus, primase does not absolutely need to form Watson−Crick hydrogen bonds to efficiently polymerize a NTP. Additionally, we found that herpes primase can misincorporate nucleotides both by misreading the template and by a primer−template slippage mechanism. The mechanistic and biological implications of these results are discussed.
Co-reporter:Kristopher E. Keller, Nisha Cavanaugh and Robert D. Kuchta
Biochemistry 2008 Volume 47(Issue 34) pp:
Publication Date(Web):August 2, 2008
DOI:10.1021/bi8008467
We analyzed the interaction of nucleoside triphosphates (NTPs) containing modified sugars to develop a better understanding of how DNA primase from herpes simplex virus I catalyzes primer synthesis. During the NTP binding reaction, primase tolerated a large number of modifications to the sugar ring. Altering the 2′ and 3′ carbons and even converting the furanose sugar into an acyclic sugar did not prevent binding. Whether or not the base on the NTP could form a correct base pair with the template base being replicated also had minimal effect on the binding reaction, indicating that primase does not use this process to discriminate between right and wrong NTPs. Rather, the key feature that primase recognizes to bind a NTP is the 5′-γ-phosphate since converting a NTP into a NDP greatly compromised binding. During the polymerization reaction, primase tolerated substantial modification of the 2′-carbon, including the presence of either an ara or ribo hydroxyl, two hydrogens, or two fluorines. However, polymerization absolutely required that the NTP contain a 3′-hydroxyl and an intact sugar ring. Modifications at the 2′-carbon of the nucleotide at the primer 3′-terminus significantly impaired further polymerization events. Compared to a ribonucleotide, incorporation of a 2′-deoxyribo- or 2′,2′-difluoro-2′-deoxyribonucleotide resulted in strong chain termination, while incorporation of an aranucleotide resulted in very strong chain termination. The implications of these data with respect to the mechanism of primase and the relationship between human and herpes primase are discussed.
Co-reporter:Julie A. Houck, Andrias Hojgaard, Joseph Piesman, Robert D. Kuchta
Ticks and Tick-borne Diseases (March 2011) Volume 2(Issue 1) pp:27-36
Publication Date(Web):1 March 2011
DOI:10.1016/j.ttbdis.2010.10.002
Lyme disease is the most common tick-borne disease in Europe and North America. In the hyperendemic Lyme disease regions of the eastern United States, nymphal Ixodes scapularis are the principal ticks transmitting the Lyme disease spirochete, Borrelia burgdorferi sensu stricto (s.s.). Approximately 25% of questing nymphs in endemic regions are infected with spirochetes. High throughput-sensitive and specific methods for testing nymphal I. scapularis for infection with B. burgdorferi are clearly needed. In the current study, we evaluated whether low-density microarrays could be adapted for the rapid and accurate detection and characterization of spirochetes in nymphal I. scapularis. Three different microarray platforms were developed and tested for the detection of spirochetes in ticks. They could both detect and differentiate different Borrelia genospecies, in one case detecting as few as a single copy of Borrelia DNA.

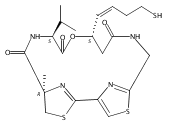
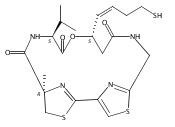
![L-Valine, N-[[(4R)-4,5-dihydro-2'-[[[(3S)-3-hydroxy-1-oxo-7-[(1-oxooctyl)thio]-4-hepten-1-yl]amino]methyl]-4-methyl[2,4'-bithiazol]-4-yl]carbonyl]-, methyl ester](/data/chemimg/191800/1415041-53-0.png)
![L-Valine, N-[[(4R)-4,5-dihydro-2'-[[[(3S)-3-hydroxy-1-oxo-7-[(1-oxooctyl)thio]-4-hepten-1-yl]amino]methyl]-4-methyl[2,4'-bithiazol]-4-yl]carbonyl]-, methyl ester](/data/chemimg/191800/1415041-53-0_b.png)




![1-Decanamine, N-[2-[[2-(4-chlorophenyl)ethyl]thio]ethyl]-, hydrochloride (1:1)](http://img.cochemist.com/data/images/1426543-79-4.png)
![1-Decanamine, N-[2-[[2-(4-chlorophenyl)ethyl]thio]ethyl]-, hydrochloride (1:1)](http://img.cochemist.com/data/images/1426543-79-4_b.png)
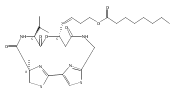
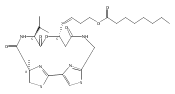
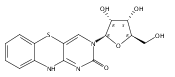
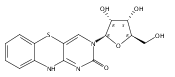
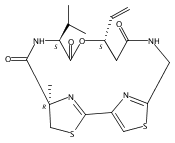
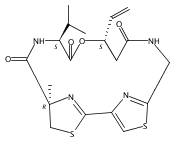

![3,7-dimethyl-3H-Imidazo[4,5-b]pyridine](http://img.cochemist.com/ccimg/893600/893566-41-1.png)
![3,7-dimethyl-3H-Imidazo[4,5-b]pyridine](http://img.cochemist.com/ccimg/893600/893566-41-1_b.png)









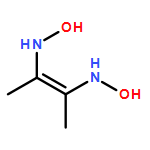
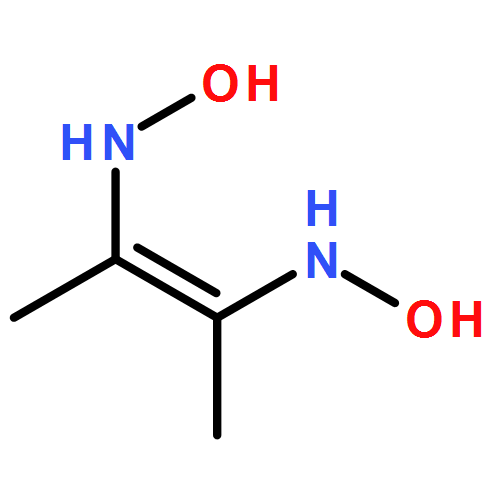

![Spiro[isobenzofuran-1(3H),9'-[9H]xanthen]-3-one,2',4',5',7'-tetrabromo-3',6'-dihydroxy-5-isothiocyanato-](http://img.cochemist.com/ccimg/60600/60520-47-0.png)
![Spiro[isobenzofuran-1(3H),9'-[9H]xanthen]-3-one,2',4',5',7'-tetrabromo-3',6'-dihydroxy-5-isothiocyanato-](http://img.cochemist.com/ccimg/60600/60520-47-0_b.png)
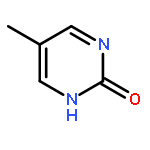
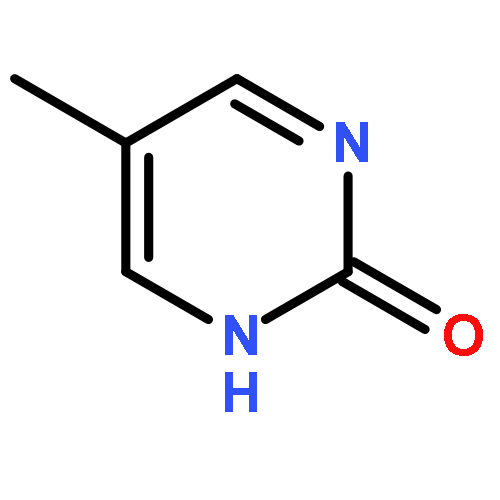


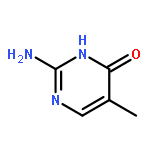

![N-[3-(4-chlorophenyl)-3,3-disulfanylpropyl]decan-1-aminium chloride](http://img.cochemist.com/ccimg/13200/13116-77-3.png)
![N-[3-(4-chlorophenyl)-3,3-disulfanylpropyl]decan-1-aminium chloride](http://img.cochemist.com/ccimg/13200/13116-77-3_b.png)


![10H-Pyrimido[5,4-b][1,4]benzothiazin-2(3H)-one](http://img.cochemist.com/ccimg/1300/1207-98-3.png)
![10H-Pyrimido[5,4-b][1,4]benzothiazin-2(3H)-one](http://img.cochemist.com/ccimg/1300/1207-98-3_b.png)