Co-reporter:Ni Tang;Wen-Xin Song;Jinyong Luo;Xiaoji Luo;Jin Chen;Katie A. Sharff;Yang Bi;Bai-Cheng He;Jia-Yi Huang;Gao-Hui Zhu;Yu-Xi Su;Wei Jiang;Min Tang;Yun He;Yi Wang;Liang Chen;Guo-Wei Zuo;Jikun Shen;Xiaochuan Pan;Russell R. Reid;Hue H. Luu;Rex C. Haydon
Journal of Cellular and Molecular Medicine 2009 Volume 13( Issue 8b) pp:2448-2464
Publication Date(Web):
DOI:10.1111/j.1582-4934.2008.00569.x
Abstract
Bone morphogenetic protein 9 (BMP-9) is a member of the transforming growth factor (TGF)-β/BMP superfamily, and we have demonstrated that it is one of the most potent BMPs to induce osteoblast differentiation of mesenchymal stem cells (MSCs). Here, we sought to investigate if canonical Wnt/β-catenin signalling plays an important role in BMP-9-induced osteogenic differentiation of MSCs. Wnt3A and BMP-9 enhanced each other’s ability to induce alkaline phosphatase (ALP) in MSCs and mouse embryonic fibroblasts (MEFs). Wnt antagonist FrzB was shown to inhibit BMP-9-induced ALP activity more effectively than Dkk1, whereas a secreted form of LPR-5 or low-density lipoprotein receptor-related protein (LRP)-6 exerted no inhibitory effect on BMP-9-induced ALP activity. β-Catenin knockdown in MSCs and MEFs diminished BMP-9-induced ALP activity, and led to a decrease in BMP-9-induced osteocalcin reporter activity and BMP-9-induced expression of late osteogenic markers. Furthermore, β-catenin knockdown or FrzB overexpression inhibited BMP-9-induced mineralization in vitro and ectopic bone formation in vivo, resulting in immature osteogenesis and the formation of chondrogenic matrix. Chromatin immunoprecipitation (ChIP) analysis indicated that BMP-9 induced recruitment of both Runx2 and β-catenin to the osteocalcin promoter. Thus, we have demonstrated that canonical Wnt signalling, possibly through interactions between β-catenin and Runx2, plays an important role in BMP-9-induced osteogenic differentiation of MSCs.
Co-reporter:Yang Bi;Jiayi Huang;Yun He;Gao-Hui Zhu;Yuxi Su;Bai-Cheng He;Jinyong Luo;Yi Wang;Quan Kang;Qing Luo;Liang Chen;Guo-Wei Zuo;Wei Jiang;Bo Liu;Qiong Shi;Min Tang;Bing-Qiang Zhang;Yaguang Weng;Ailong Huang;Lan Zhou;Tao Feng;Hue H. Luu;Rex C. Haydon;Ni Tang
Journal of Cellular Biochemistry 2009 Volume 108( Issue 1) pp:295-303
Publication Date(Web):
DOI:10.1002/jcb.22254
Abstract
Wnt/β-catenin pathway plays an important role in regulating embryonic development. Hepatocytes differentiate from endoderm during development. Hepatic progenitor cells (HPCs) have been isolated from fetal liver and extrahepatic tissues. Most current studies in liver development and hepatic differentiation have been focused on Wnts, β-catenin, and their receptors. Here, we sought to determine the role of Wnt antagonists in regulating hepatic differentiation of fetal liver-derived HPCs. Using mouse liver tissues derived from embryonic day E12.5 to postnatal day (PD) 28, we found that 13 of the 19 Wnt genes and almost all of Wnt receptors/co-receptors were expressed in most stages. However, Wnt antagonists SFRP2, SFRP3, and Dkk2 were only detected in the early stages. We established and characterized the reversible stable HPCs derived from E14.5 mouse fetal liver (HP14.5). HP14.5 cells were shown to express high levels of early liver progenitor cell markers, but low levels or none of late liver markers. HP14.5 cells were shown to differentiate into mature hepatocytes upon dexamethasone (Dex) stimulation. Dex-induced late marker expression and albumin promoter activity in HP14.5 cells were inhibited by exogenous expression of SFRP3. Furthermore, Dex-induced glycogen synthesis of PAS-positive HP14.5 cells was significantly inhibited by SFRP3. Therefore, our results have demonstrated that the expression of Wnt antagonists decreases as hepatic differentiation progresses, suggesting that a balanced Wnt signaling may be critical during mouse liver development and hepatic differentiation. J. Cell. Biochem. 108: 295–303, 2009. © 2009 Wiley-Liss, Inc.
Co-reporter:Xiaoji Luo, Jin Chen, Wen-Xin Song, Ni Tang, Jinyong Luo, Zhong-Liang Deng, Katie A Sharff, Gary He, Yang Bi, Bai-Cheng He, Erwin Bennett, Jiayi Huang, Quan Kang, Wei Jiang, Yuxi Su, Gao-Hui Zhu, Hong Yin, Yun He, Yi Wang, Jeffrey S Souris, Liang Chen, Guo-Wei Zuo, Anthony G Montag, Russell R Reid, Rex C Haydon, Hue H Luu and Tong-Chuan He
Laboratory Investigation 2008 88(12) pp:1264-1277
Publication Date(Web):October 6, 2008
DOI:10.1038/labinvest.2008.98
Osteosarcoma (OS) is the most common primary malignancy of bone. Here, we investigated a possible role of defective osteoblast differentiation in OS tumorigenesis. We found that basal levels of the early osteogenic marker alkaline phosphatase (ALP) activity were low in OS lines. Osteogenic regulators Runx2 and OSX, and the late marker osteopontin (OPN) expressed at low levels in most OS lines, indicating that most OS cells fail to undergo terminal differentiation. Furthermore, OS cells were refractory to osteogenic BMP-induced increases in ALP activity. Osteogenic BMPs were shown to upregulate early target genes, but not late osteogenic markers OPN and osteocalcin (OC). Furthermore, osteogenic BMPs failed to induce bone formation from human OS cells, rather effectively promoted OS tumor growth in an orthotopic OS model. Exogenous expression of early target genes enhanced BMP-stimulated OS tumor growth, whereas osteogenic BMP-promoted OS tumor growth was inhibited by exogenous Runx2 expression. These results suggest that alterations in osteoprogenitors may disrupt osteogenic differentiation pathway. Thus, identifying potential differentiation defects in OS tumors would allow us to reconstruct the tumorigenic events in osteoprogenitors and to develop rational differentiation therapies for clinical OS management.
Co-reporter:
Nature Protocols 2007 2(5) pp:
Publication Date(Web):2007-05-17
DOI:10.1038/nprot.2007.135
The most commonly used adenoviral vectors are derived from human adenovirus serotypes 2 and 5. Adenoviruses are non-enveloped DNA viruses whose capsid is composed of pentons and hexons. The viral genome consists of 36-kb double-stranded linear DNA with inverted terminal repeat (ITR) sequences at each end. DNA of length greater than 38 kb cannot be efficiently packaged into competent viral particles. The adenovirus life cycle begins with the attachment of the fiber to cell surface receptors, such as coxsackievirus and adenovirus receptor, and the interaction of the pentons with αvβ3 and αvβ5 integrin proteins. Following receptor-mediated endocytosis, adenovirus escapes from the endosomes to the cytoplasm and translocates into the nucleus, where viral transcription and replication begin. Completion of the virus life cycle triggers cell death and the release of progeny viruses. On the basis of temporal expression relative to the onset of viral DNA replication, viral transcription units are conventionally referred to as early (E1a, E1b, E2, E3 and E4), delayed early (proteins IX and Iva2) or late genes (L1–L5). The early gene products are generally involved in viral gene transcription, DNA replication, host immune suppression and inhibition of host cell apoptosis, whereas the late gene products are required for virion assembly8.The generation and production of recombinant adenoviruses should be performed in a laboratory operating at Biosafety Level 2 (BL2), as approved by the researcher's institutional biosafety committee and following the National Institutes of Health Biosafety guidelines (http://www.cdc.gov/od/ohs/biosfty/bmbl4/bmbl4toc.htm). These requirements include the use of biosafety cabinet hoods, the establishment of proper procedures for decontamination and disposal of liquid and solid wastes and the disinfection of contaminated surfaces and equipment. In addition, users should conduct a regular analysis of adenovirus stocks to exclude the presence of replication-competent adenoviruses in their preparations.The AdEasy system described in this protocol exploits the high efficiency of homologous recombination in a specific bacterial strain coupled with selectable antibiotic resistance markers to simplify recombinant vector production4, 7, 20.Two adenoviral backbone vectors can be used for adenovirus production (Fig. 2b). The commonly used pAdEasy-1 is an E1 and E3 double-deletion vector. Hence, AdEasy-1-derived recombinant adenoviruses can be propagated in E1-expressing packaging cells, such as HEK-293 cells (E3 is not necessary for viral production). The pAdEasy-2 backbone vector is an E1, E3 and E4 triple-deletion vector7, 20. Thus, propagation of AdEasy-2-derived vectors requires the use of packaging cell lines expressing both E1 and E4. As the E4 gene product is toxic to mammalian cells, its expression is usually controlled by inducible promoters. Several such E1/E4 packaging lines are available. However, because of the leakiness of most inducible systems, high E4-expressing cells are often lost after serial passages, and it is difficult to generate high-titer AdEasy-2-derived viruses. Thus, in this protocol we focus on procedures for generating recombinant adenoviruses with the AdEasy-1 system. For investigators wishing to express particularly long (or multiple) transgenes, we recommend gutless systems. Unlike AdEasy vector systems, which can tolerate replacements in the E1 and E4 regions only, gutless systems can tolerate much larger replacements in the adenoviral backbone4, 5, 6, 11. Although the AdEasy system has been made freely available to thousands of academic researchers, the system can also be obtained through commercial sources listed on the AdEasy website (http://www.coloncancer.org/adeasy.htm).
Note: As BJ5183 cells have a relatively high frequency of homologous recombination, unwanted or detrimental rearrangements and/or recombinations of the pAdEasy sequences in AdEasier cells can occur. For homemade AdEasier cells, it is thus important to pick individual clones and characterize the clones with extensive restriction digestions, for example with HindIII and/or PstI. The digestion patterns can be compared with the pAdEasy stock DNA made in a strain incapable of homologous recombination (e.g., DH10B or XL-1 blue).
Note: All shuttle vectors confer resistance to kanamycin.
Note: All cell culture work involved in adenovirus generation and subsequent amplification should be performed in accordance with BL2 guidelines.
Note: Special precautions are needed when washing HEK-293 cells because the cells are often weakly adherent to the flask. One wash is usually sufficient. If multiple flasks are used for transfections, wash no more than five flasks at a time to minimize detachment of HEK-293 cells.
Note: The transfected cells (and the subsequent cell cultures) should be treated as a BL2 biohazard and handled with care, as the medium may contain recombinant adenoviruses.
Note: No obvious plaques or cytopathic effect (CPE) is generally observed by light microscopy up to 2 weeks after transfection. However, GFP plaques are usually observed under fluorescence microscopy from 5–7 d after transfection (Fig. 4a). The shapes of the fluorescent plaques depend on transfection efficiency. Low transfection efficiencies may produce 'comet-like' plaques, whereas high transfection efficiencies may produce an intense 'scattered stars' phenomenon.
Note: We have generally been able to generate adenoviruses suitable for experimental use in tissue culture without plaque purification. For more exacting applications, however, it is advisable to perform plaque purification. Homogeneity of adenoviruses can be judged by restriction endonuclease digestion, paying careful attention to the ratio of the fragments obtained (which should be molar if the viruses are homogeneous). We have not seen evidence of recombination within more than 50 adenoviruses produced with the AdEasy system. However, we cannot exclude subtle changes in sequence that may occur during the cloning or viral production steps. If such subtle changes are an issue, the GOI can be amplified from the final viral preparation using PCR and the sequence determined using conventional means.
Note: Perform infection under BL2 conditions.
Note: The virus-containing waste should be disinfected with chlorine bleach.
Note: Titers can be measured at any time, which is particularly easy with AdTrack-based vectors. Simply infect HEK-293 cells with various dilutions of viral supernatant and see how many are GFP-positive cells 24 h later. Without AdTrack, viruses can be plaque titered or titered by limiting dilution using standard methods7. After three rounds of amplification, viral titer should reach 109–1010 infectious particles [or plaque-forming units (pfu)] per milliliter of lysate.
Note: The virus-containing waste should be disinfected with chlorine bleach.Step 1, cloning GOI into a shuttle vector: 3–7 dSteps 2–12, generating recombinant adenovirus plasmids: 4–5 dSteps 13–20, generating the initial stocks of adenoviruses: 14–20 dSteps 21–29, stepwise amplification for high-titer adenoviruses: 7–10 dStep 31, determining viral titers: 2–10 dTroubleshooting advice can be found in Table 1.The AdEasy system has proved to be an efficient and robust technology for generating recombinant adenoviruses in thousands of laboratories worldwide. The key step is the generation of recombinants in AdEasier cells. Once these recombinants are obtained, generation of adenoviruses in HEK-293 cells is virtually guaranteed23, 24, 25, 26, 27. Depending on the transfection efficiency of HEK-293 cells, one can expect titers in the initial lysate to be 108– 1010 pfu ml−1. It usually requires two to three rounds of further amplification in HEK-293 cells to achieve viral titers of 1012 or 1013 pfu ml−1. Cloning genes of interest into shuttle vectors entails standard molecular biology techniques. The user's experience in molecular cloning will determine the time required for this step.After subcloning GOI into a shuttle vector, the rate-limiting step is generally the production of the desired recombinants in AdEasier cells. With experience and high-quality competent AdEasier cells, it takes as few as 2 d to obtain and verify the recombinants.If every step works well, one can expect to obtain the initial virus lysates 2–3 weeks thereafter and the subsequent large-scale virus preparation in an additional 1–2 weeks (see TIMING). Further information can be found at the Internet resources listed in Box 1.
Co-reporter:Jinyong Luo, Jin Chen, Zhong-Liang Deng, Xiaoji Luo, Wen-Xin Song, Katie A Sharff, Ni Tang, Rex C Haydon, Hue H Luu and Tong-Chuan He
Laboratory Investigation 2007 87(2) pp:97-103
Publication Date(Web):January 8, 2007
DOI:10.1038/labinvest.3700509
Wnt signaling plays an important role in regulating cell proliferation and differentiation. De-regulation of these signaling pathways has been implicated in many human diseases, ranging from cancers to skeletal disorders. Wnt proteins are a large family of secreted factors that bind to the Frizzled receptors and LRP5/6 co-receptors and initiate complex signaling cascades. Over the past two decades, our understanding of Wnt signaling has been significantly improved due to the identification of many key regulators and mediators of these pathways. Given that Wnt signaling is tightly regulated at multiple cellular levels, these pathways themselves offer ample nodal points for targeted therapeutics. Here, we focus on our current understanding of these pathways, the associations of Wnt signaling with human disorders, and the opportunities to target key components of Wnt signaling for rational drug discovery.
Co-reporter:Quan Kang;Michael H. Sun;Xinmin Li;Jae Yoon Park;Rex C. Haydon;Hongwei Cheng;Ying Peng;Hue H. Luu;Wei Jiang
Journal of Cellular Biochemistry 2003 Volume 90(Issue 6) pp:1149-1165
Publication Date(Web):5 NOV 2003
DOI:10.1002/jcb.10744
Bone formation is presumably a complex and well-orchestrated process of osteoblast lineage-specific differentiation. As members of the TGFβ superfamily, bone morphogenetic proteins (BMPs) play an important role in regulating osteoblast differentiation and subsequent bone formation. Several BMPs are able to induce de novo bone formation. Although significant progress has recently been made about the transcriptional control of osteoblast differentiation, detailed molecular events underlying the osteogenic process remain to be elucidated. In order to identify potentially important signaling mediators activated by osteogenic BMPs but not by non-osteogenic BMPs, we sought to determine the transcriptional differences between three osteogenic BMPs (i.e., BMP-2, BMP-6, and BMP-9) and two inhibitory/non-osteogenic BMPs (i.e., BMP-3 and BMP-12). Through the microarray analysis of approximately 12,000 genes in pre-osteoblast progenitor cells, we found that expression level of 203 genes (105 up-regulated and 98 down-regulated) was altered >2-fold upon osteogenic BMP stimulation. Gene ontology analysis revealed that osteogenic BMPs, but not inhibitory/non-osteogenic BMPs, activate genes involved in the proliferation of pre-osteoblast progenitor cells towards osteoblastic differentiation, and simultaneously inhibit myoblast-specific gene expression. BMP-regulated expression of the selected target genes was confirmed by RT-PCR, as well as by the CodeLink Bioarray analysis. Our findings are consistent with the notion that osteogenesis and myogenesis are two divergent processes. Further functional characterization of these downstream target genes should provide important insights into the molecular mechanisms behind BMP-mediated bone formation. © 2003 Wiley-Liss, Inc.
Co-reporter:Ke Yang, Jin Chen, Wei Jiang, Enyi Huang, ... Tong-Chuan He
Journal of Investigative Dermatology (October 2012) Volume 132(Issue 10) pp:2479-2483
Publication Date(Web):1 October 2012
DOI:10.1038/jid.2012.145
Co-reporter:Qing Luo, Quan Kang, Wen-Xin Song, Hue H. Luu, Xiaoji Luo, Naili An, Jinyong Luo, Zhong-Liang Deng, Wei Jiang, Hong Yin, Jin Chen, Katie A. Sharff, Ni Tang, Erwin Bennett, Rex C. Haydon, Tong-Chuan He
Gene (15 June 2007) Volume 395(Issues 1–2) pp:160-169
Publication Date(Web):15 June 2007
DOI:10.1016/j.gene.2007.02.030
RNA interference (RNAi)-mediated gene silencing has become a valuable tool for functional studies, reverse genomics, and drug discoveries. One major challenge of using RNAi is to identify the most effective short interfering RNAs (siRNAs) sites of a given gene. Although several published bioinformatic prediction models have proven useful, the process to select and validate optimal siRNA sites for a given gene remains empirical and laborious. Here, we developed a fluorescence-based selection system using a retroviral vector backbone, namely pSOS, which was based on the premise that candidate siRNAs would knockdown the chimeric transcript between GFP and target gene. The expression of siRNA was driven by the opposing convergent H1 and U6 promoters. This configuration simplifies the cloning of duplex siRNA oligonucleotide cassettes. We demonstrated that GFP signal reduction was closely correlated with siRNA knockdown efficiency of human β-catenin, as well as with the inhibition of β-catenin/Tcf4 signaling activity. The pSOS should not only facilitate the selection and validation of candidate siRNA sites, but also provide efficient delivery tools of siRNAs via viral vectors in mammalian cells. Thus, the pSOS system represents an efficient and user-friendly strategy to select and validate siRNA target sites.
.jpg)
![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8.png)
![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8_b.png)


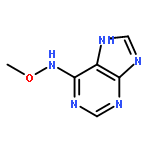
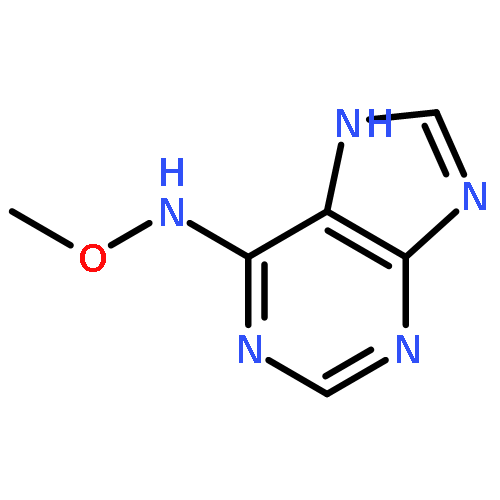

















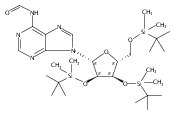
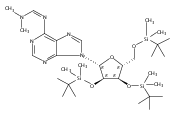






































































































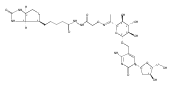




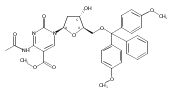



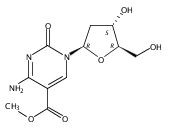
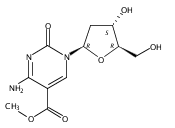










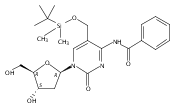


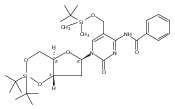
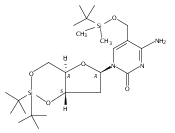

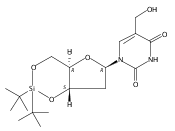



![1H-Thieno[3,4-d]imidazole-4-pentanamide, hexahydro-2-oxo-N-3,6,9,12-tetraoxapentadec-14-yn-1-yl-, (3aS,4S,6aR)-](/data/chemimg/186200/1262681-31-1.png)
![1H-Thieno[3,4-d]imidazole-4-pentanamide, hexahydro-2-oxo-N-3,6,9,12-tetraoxapentadec-14-yn-1-yl-, (3aS,4S,6aR)-](/data/chemimg/186200/1262681-31-1_b.png)





















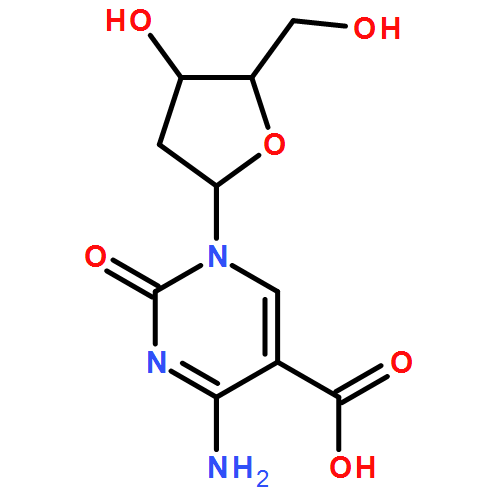


![5,8,11-Trioxa-2,14-diazanonadecanoic acid, 19-[(3aS,4S,6aR)-hexahydro-2-oxo-1H-thieno[3,4-d]imidazol-4-yl]-15-oxo-, 11,12-didehydro-5,6-dihydrodibenzo[a,e]cycloocten-5-yl ester](/data/chemimg/186100/1027338-10-8.png)
![5,8,11-Trioxa-2,14-diazanonadecanoic acid, 19-[(3aS,4S,6aR)-hexahydro-2-oxo-1H-thieno[3,4-d]imidazol-4-yl]-15-oxo-, 11,12-didehydro-5,6-dihydrodibenzo[a,e]cycloocten-5-yl ester](/data/chemimg/186100/1027338-10-8_b.png)







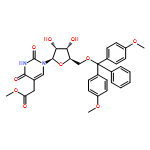






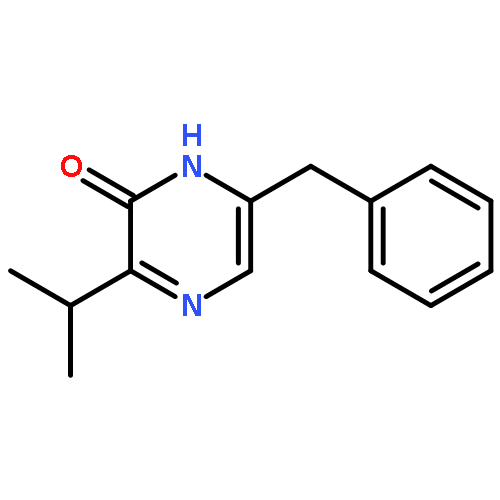




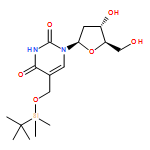










![1H-Thieno[3,4-d]imidazole-4-pentanoicacid, hexahydro-2-oxo-, 2-[2-(aminooxy)acetyl]hydrazide, (3aS,4S,6aR)-](http://img.cochemist.com/ccimg/139600/139585-03-8.png)
![1H-Thieno[3,4-d]imidazole-4-pentanoicacid, hexahydro-2-oxo-, 2-[2-(aminooxy)acetyl]hydrazide, (3aS,4S,6aR)-](http://img.cochemist.com/ccimg/139600/139585-03-8_b.png)