Co-reporter:Mumtaz Kasim, Huai-Chun Chen and Richard P. Swenson
Biochemistry 2009 Volume 48(Issue 23) pp:
Publication Date(Web):May 8, 2009
DOI:10.1021/bi900607q
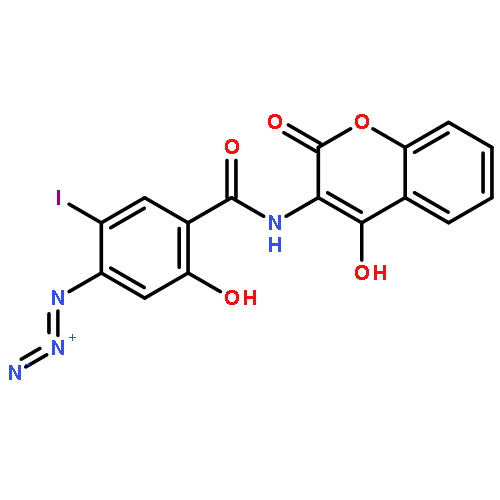
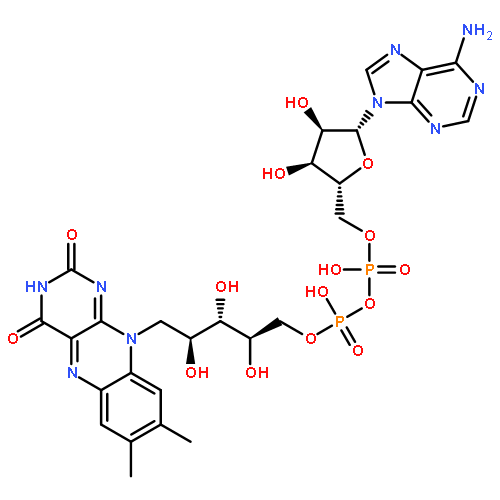
Flavocytochrome P450BM-3, a bacterial monooxygenase, contains a flavin mononucleotide-binding domain bearing a strong structural homology to the bacterial flavodoxin. The flavin mononucleotide (FMN) serves as the one-electron donor to the heme iron, but in contrast to the electron transfer mechanism of mammalian cytochrome P450 reductase, the FMN semiquinone state is not thermodynamically stable and appears transiently as the anionic rather than the neutral form. A unique loop region comprised of residues 536Y-N-G-H-P-P541, which forms a type I′ reverse turn and provides several interactions with the FMN isoalloxazine ring, was targeted in this study. Nuclear magnetic resonance studies support the presence of a strong hydrogen bond between the backbone amide of Asn537 and FMN N5, the anionic ionization state of the hydroquinone, and for a change in the hybridization state of the N5 upon reduction. Replacement of Tyr536, which flanks the flavin ring, with a basic residue (histidine or arginine) did not significantly influence the redox properties of the FMN or the accumulation of the anionic semiquinone. The central residues of the type I′ turn (Asn-Gly) were replaced with various combinations of glycine and alanine as a means of altering the turn and its interactions. Gly538 was found to be crucial in maintaining the type I′ turn conformation of the loop and the strong H-bonding interaction at N5. The functional role of the tandem Pro-Pro sequence which anchors and possible “rigidifies” the loop was investigated through alanine replacements. Despite changes in the stabilities of the oxidized and hydroquinone redox states of the FMN, none of the replacements studied significantly altered the two-electron midpoint potentials. Pro541 does contribute to some degree to the strength of the N5 interaction and the formation of the anionic semiquinone. Unlike that of the flavodoxin, it would appear that the conformation of the FMN rather than the loop changes in response to reduction in this flavoprotein.
Co-reporter:Huai-Chun Chen and Richard P. Swenson
Biochemistry 2008 Volume 47(Issue 52) pp:13788-13799
Publication Date(Web):December 4, 2008
DOI:10.1021/bi800954h
Despite sharing sequence and structural similarities with other diflavin reductases such as NADPH-cytochrome P450 reductase (CPR) and nitric oxide synthase, flavocytochrome P450BM-3 displays some unique redox and electron transferring properties, including the inability to thermodynamically stabilize the neutral semiquinone (SQ) state of the flavin mononucleotide (FMN) cofactor. Rather, the anionic SQ species is only transiently formed during rapid reduction. Why is this? The absence of a conserved glycine residue and, as a consequence, the shorter and less flexible cofactor-binding loop in P450BM-3 represents a notable difference from other diflavin reductases and the structurally related flavodoxin. This difference may facilitate the formation of a strong hydrogen bond between backbone amide NH group of Asn537 and N5 of the oxidized FMN, an interaction not found in the other proteins. In the flavodoxin, the conserved glycine residue plays a crucial role in a redox-linked conformational change that contributes to the thermodynamic stabilization of the neutral SQ species of the FMN through the formation of a hydrogen bond with the N5H group of the flavin. In this study, a glycine residue was inserted after Tyr536 in the loop within the isolated FMN-binding domain as well as the diflavin reductase domain of P450BM-3, a position equivalent to Gly141 in human CPR. As a result, the insertion variant was observed to accumulate the neutral form of the FMN SQ species much like CPR. The midpoint potential for the SQ/HQ couple decreased by 68 mV, while that for the OX/SQ couple remained unchanged. 15N NMR data provide evidence of the disruption of the hydrogen bond between the backbone amide group of Asn537 and the N5 atom in the oxidized state of the FMN. Molecular models suggest that the neutral FMN SQ could be stabilized through hydrogen bonding with the backbone carbonyl group of the inserted glycine residue in a manner similar to that of CPR and the flavodoxin. The insertion of the glycine at the same location within the diflavin domain resulted in a purified protein that retained nearly stoichiometric levels of bound FAD but tended to lose the FMN cofactor. This preparation retained one-third of the ferricyanide reductase activity but <1% of the cytochrome c reductase activity of the wild type. However, the insertion variant reconstituted with FMN regained nearly half of the wild-type cytochrome c reductase activity. These results demonstrate the importance of the unique structural characteristics of the shorter loop in P450BM-3 in establishing the unique redox properties of the FMN in this protein but not its general cytochrome reductase activity.
.jpg)

![2,5-Pyrrolidinedione, 1-[(4-azido-2-hydroxy-5-iodobenzoyl)oxy]-](http://img.cochemist.com/ccimg/99300/99298-06-3.png)
![2,5-Pyrrolidinedione, 1-[(4-azido-2-hydroxy-5-iodobenzoyl)oxy]-](http://img.cochemist.com/ccimg/99300/99298-06-3_b.png)



![Acetamide, 2,2,2-trifluoro-N-[6-(phosphonooxy)hexyl]-](http://img.cochemist.com/ccimg/40300/40248-29-1.png)
![Acetamide, 2,2,2-trifluoro-N-[6-(phosphonooxy)hexyl]-](http://img.cochemist.com/ccimg/40300/40248-29-1_b.png)




![Benzo[g]pteridine-2,4(1H,3H)-dione](http://img.cochemist.com/ccimg/500/490-59-5.png)
![Benzo[g]pteridine-2,4(1H,3H)-dione](http://img.cochemist.com/ccimg/500/490-59-5_b.png)


![3-[(4-AZIDOPHENYL)METHYL]-4-HYDROXYCHROMEN-2-ONE](http://img.cochemist.com/ccimg/134000/133950-79-5.png)
![3-[(4-AZIDOPHENYL)METHYL]-4-HYDROXYCHROMEN-2-ONE](http://img.cochemist.com/ccimg/134000/133950-79-5_b.png)