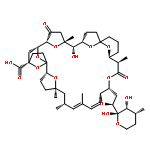
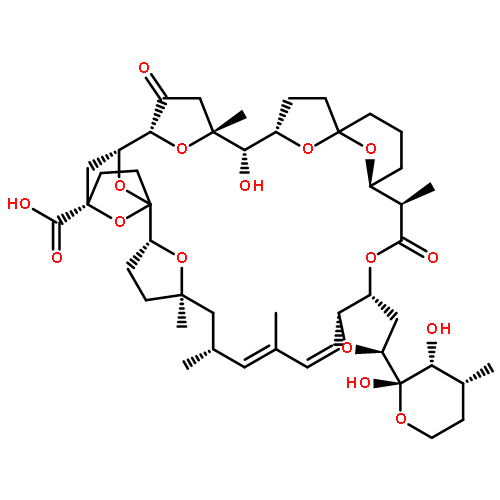
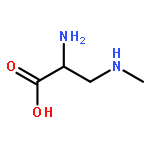
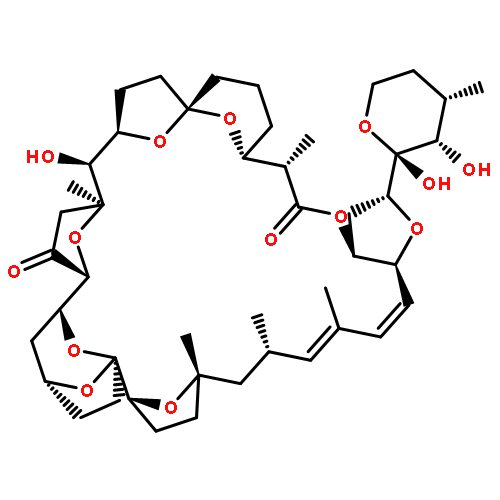
Co-reporter:Ling Ding, Jiangbing Qiu, and Aifeng Li
Journal of Agricultural and Food Chemistry July 12, 2017 Volume 65(Issue 27) pp:5494-5494
Publication Date(Web):June 15, 2017
DOI:10.1021/acs.jafc.7b02101
A seafood poisoning event occurred in Qinhuangdao, China, in April 2016. Subsequently, the causative mussels (Mytilus galloprovincialis) were harvested and analyzed to reveal a high concentration [∼10 758 μg of saxitoxin (STX) equiv kg–1] of paralytic shellfish toxins (PSTs), including gonyautoxin (GTX)1/4 and GTX2/3, as well as new metabolites 11-hydroxy-STX (M2), 11,11-dihydroxy-STX (M4), open-ring 11,11-dihydroxy-STX (M6), 11-hydroxy-neosaxitoxin (NEO) (M8), and 11,11-dihydroxy-NEO (M10). To understand the origin and biotransformation pathways of these new metabolites, uncontaminated mussels (M. galloprovincialis) were fed with either of two Alexandrium tamarense strains (ATHK and TIO108) under laboratory conditions. Similar PST metabolites were also detected in mussels from both feeding experiments. Results supposed that 11-hydroxy-C2 toxin (M1) and 11,11-dihydroxy-C2 (M3) are transformed from C2, while 11-hydroxy-C4 toxin (M7) and 11,11-dihydroxy-C4 (M9) are converted from C4. In addition, the metabolites M2, M4, and M6 appear to be products of GTX2/3, and the metabolites M8 and M10 are likely derived from GTX1/4.Keywords: Alexandrium tamarense; biotransformation; metabolites; Mytilus galloprovincialis; paralytic shellfish toxins (PSTs);
Co-reporter:Meihui Li, Geng Sun, Jiangbing Qiu, Aifeng Li
Toxicon 2017 Volume 127(Volume 127) pp:
Publication Date(Web):1 March 2017
DOI:10.1016/j.toxicon.2016.12.009
•Lipophilic shellfish toxins were monitored in coastal aquaculture zone of Qingdao.•Microalgae, scallop, and seawater samples were collected by multiple approaches.•Dinophysis acuminata, D. fortii, and Phalacroma rotundatum were identified here.•A novel compound isobaric DTX1 was found only in SPATT bags deployed in field.•A discrepancy was mentioned for OA and DTX1 adsorbed by the SPATT bags in field.Lipophilic shellfish toxins (LSTs) produced by diverse microalgae have become a potential threat to the marine aquaculture industry in coastal regions throughout the world. In this study, phytoplankton, scallop, and seawater samples were collected and monitored in the aquaculture zone of Liuqing Bay located in the coast of Qingdao, Yellow Sea, in 2014 and 2015. Dissolved LSTs in seawater were adsorbed by solid phase adsorption toxin tracking (SPATT) bags and solid phase extraction (SPE) cartridges, respectively. PTX2 was found to be the predominant LSTs, while OA and DTX1 were detected at trace levels in the three species of Dinophysis and Phalacroma genera picked from the investigation zone. The highest quota of PTX2 occurred in D. fortii (0.28 pg cell−1), followed by D. acuminata complex (0.08 pg cell−1) and P. rotundatum (=D. rotundata) (0.02 pg cell−1). PTX2 (nd∼5.7 μg kg−1), OA (nd∼2.8 μg kg−1) and DTX1 (nd∼1.6 μg kg−1) were also observed in cultivated scallops, but in addition to the former, PTX2sa, 7-epi-PTX2sa and an isobaric form of DTX1 suspected as DTX1b were also found in the SPATT bags. The isobaric compound of DTX1 only appeared in the SPATT bags deployed in field samples of seawater. A significant higher quantity of OA was adsorbed by SPATT bags compared to DTX1, although similar concentrations of both toxins were adsorbed by SPE cartridges from the aquaculture zone. AZA2 was also found in the concentrated phytoplankton cells, which demonstrated that AZA-producing microalgae were present in this investigated area. The variation in concentrations of PTX2 adsorbed by SPATT bags could reflect the change of Dinophysis density in seawater, but there was a time lag behind the peaks of Dinophysis density. Comparative results did not support the SPATT technology as a useful warning tool for toxic algal blooms and toxin accumulation in bivalves. The data suggest that the contamination of LSTs was prevalent in the coast of Qingdao and the environmental behavior of LSTs should be explored further.Download high-res image (393KB)Download full-size image
Co-reporter:Lin Fan, Geng Sun, Jiangbing Qiu, Qimin Ma, Philipp Hess, Aifeng Li
Journal of Chromatography A 2014 Volume 1373() pp:1-8
Publication Date(Web):19 December 2014
DOI:10.1016/j.chroma.2014.11.008
•HP20 resin bags were deployed in three artificial seawaters at different salinity.•Dynamic adsorption behavior of OA and DTX1 by these SPATT bags was explored.•The highest initial sorption rate of toxins occurred in seawater at medium salinity.•Resin pores below 10 nm in size governed adsorption in natural seawater.•Toxins were retained by pores in seawaters at high and low salinity.In the present study, okadaic acid (OA) and dinophysistoxin-1 (DTX1) were spiked into artificial seawater at low, medium and high estuarine salinities (9‰, 13.5‰ and 27‰). Passive samplers (HP20 resin) used for solid phase adsorption toxin tracking (SPATT) technology were exposed in these seawaters for 12-h periods. Adsorption curves well fitted a pseudo-secondary kinetics model. The highest initial sorption rates of both toxins occurred in the seawater of medium salinity, followed by seawater of low and high estuarine salinity. Pore volumes of micropores (<2 nm) and small mesopores (2 nm < diameter < 10 nm) of HP20 resin decreased after adsorption of toxins in seawater at high and low salinity but not in seawater at medium salinity, which demonstrated that the toxin molecules entered into micropores and mesopores (below 10 nm in size) in seawaters of high and low salinity. More toxin or other matrix agglomerates were displayed on the surface of resin deployed in the seawater of medium salinity. Taking into consideration the pore-size distribution and surface images, it appears that intra-particle diffusion governs toxin adsorption in seawater at high salinity while film diffusion mainly controls the adsorption process in seawater at medium salinity. This is the first study to confirm that molecules of OA and DTX1 are able to enter into micropores (<2 nm) and small mesopores (2–10 nm) of HP20 resin in estuarine seawater with high salinity (∼27‰).
Co-reporter:Hui Zhao, Jiangbing Qiu, Hua Fan, Aifeng Li
Journal of Chromatography A 2013 1300() pp: 159-164
Publication Date(Web):
DOI:10.1016/j.chroma.2013.02.048
Co-reporter:Aifeng Li, Feifei Ma, Xiuli Song, Rencheng Yu
Journal of Chromatography A 2011 Volume 1218(Issue 11) pp:1437-1442
Publication Date(Web):18 March 2011
DOI:10.1016/j.chroma.2011.01.043
Solid-phase adsorption toxin tracking (SPATT) technology was developed as an effective passive sampling method for dissolved diarrhetic shellfish poisoning (DSP) toxins in seawater. HP20 and SP700 resins have been reported as preferred adsorption substrates for lipophilic algal toxins and are recommended for use in SPATT testing. However, information on the mechanism of passive adsorption by these polymeric resins is still limited. Described herein is a study on the adsorption of OA and DTX1 toxins extracted from Prorocentrum lima algae by HP20 and SP700 resins. The pore size distribution of the adsorbents was characterized by a nitrogen adsorption method to determine the relationship between adsorption and resin porosity. The Freundlich equation constant showed that the difference in adsorption capacity for OA and DTX1 toxins was not determined by specific surface area, but by the pore size distribution in particular, with micropores playing an especially important role. Additionally, it was found that differences in affinity between OA and DTX1 for aromatic resins were as a result of polarity discrepancies due to DTX1 having an additional methyl moiety.
Co-reporter:Aifeng Li, Huidan Chen, Jiangbing Qiu, Heshan Lin, Haifeng Gu
Toxicon (January 2016) Volume 109() pp:84-93
Publication Date(Web):1 January 2016
DOI:10.1016/j.toxicon.2015.11.016
•Mollusks collected from the Chukchi and Bering seas were analyzed using LC-MS/MS.•PTX2 and STX were detected exclusively in mollusks sampled from the Chukchi Sea.•OA and DTX1 were determined regionally in mollusks originated in the Bering Sea.•SPX1, YTX, and some analogues occurred in most of samples from both sampling sites.•No Azaspiracid 1–3, Gymnodimine and Pinnatoxin G were found in any samples tested.Buccinidae whelk Neptunea varicifera (Dall), Cardiidae clam Serripes laperousii (Deshayes), and two unknown species of whelk and clam were collected from the Arctic Chukchi Sea and sub-Arctic Bering Sea in July 2014. In this study, the mollusk samples were analyzed by different liquid chromatography-tandem quadrupole mass spectrometry (LC-MS/MS) methods for multiple shellfish toxins, including okadaic acid (OA), pectenotoxin (PTX), yessotoxin (YTX), azaspiracid (AZA), cyclic imines (CI), and saxitoxin (STX) groups. PTX2 (≈2.0 μg kg−1 whole tissues) was detected exclusively in the clam S. laperousii collected from the Chukchi Sea. OA and dinophysistoxin-1 (DTX1) were restricted to mollusk samples collected from the Bering Sea, and OA was the dominant component of the whelk N. varicifera (63 μg kg−1 digestive gland) and an unknown species of whelk (6.8 μg kg−1 digestive gland). Spirolide-1 (SPX1) was confirmed in most samples except for the whelk N. varicifera collected from the Bering Sea. The highest content of SPX1 (≈18.5 μg kg−1 digestive gland) occurred in the whelk N. varicifera collected from the Chukchi Sea, along with the suspected presence of SPX-C, SPX-D and didesMe-SPX-C. YTX, as well as its derivatives 45-OH-YTX and 45,46,47-Trinor-YTX, were found in all samples, with the highest YTX content (66 μg kg−1 digestive gland) present in the whelk N. varicifera collected from the Chukchi Sea. Interestingly, STX and dcSTX were measured only in the whelk N. varicifera and unknown species of clam collected from the Chukchi Sea. No AZA-group toxins, gymnodimine (GYM), or pinnatoxin G were found in any samples analyzed. Results demonstrated that the mollusk samples were contaminated by multiple shellfish toxins in the Chukchi and Bering seas. This study highlights the need to monitor potentially toxic microalgae in the Arctic and sub-Arctic regions, as well as species of mollusk that may be included in future commercial or subsistence harvests.
Co-reporter:Aifeng Li, Jinggang Ma, Jijuan Cao, Pearse McCarron
Toxicon (1 September 2012) Volume 60(Issue 3) pp:420-425
Publication Date(Web):1 September 2012
DOI:10.1016/j.toxicon.2012.04.339
More than 200 people in China suffered illness with symptoms of diarrhetic shellfish poisoning (DSP) following consumption of mussels (Mytilus galloprovincialis). The event occurred in the cities of Ningbo and Ningde near the East China Sea in May, 2011. LC–MS/MS analysis showed that high concentrations of okadaic acid, dinophysistoxin-1, and their acyl esters were responsible for the incidents. The total concentration was more than 40 times the EU regulatory limit of 160 μg OA eq./kg. Pectentoxin-2 and its seco-acids were also present in the mussels. Additionally, yessotoxins were found to be responsible for positive mouse bioassay results on scallop (Patinopecten yessoensis) and oyster (Crassostrea talienwhanensis) samples collected from the North Yellow Sea in June, 2010.This work shows that high levels of lipophilic toxins can accumulate in shellfish from the Chinese coast and it emphasises that adequate chemical analytical methodologies are needed for monitoring purposes. Further research is required to broaden the knowledge on the occurrence of lipophilic toxins in Chinese shellfish.
Co-reporter:Jiangbing Qiu, Feifei Ma, Hua Fan, Aifeng Li
Aquaculture (1 June 2013) Volumes 396–399() pp:76-81
Publication Date(Web):1 June 2013
DOI:10.1016/j.aquaculture.2013.02.040

![1H,10H-Pyrrolo[1,2-c]purine-10,10-diol,2,6-diamino-4-[[(aminocarbonyl)oxy]methyl]-3a,4,8,9-tetrahydro-, (3aS,4R,10aS)-](http://img.cochemist.com/ccimg/35600/35523-89-8.png)
![1H,10H-Pyrrolo[1,2-c]purine-10,10-diol,2,6-diamino-4-[[(aminocarbonyl)oxy]methyl]-3a,4,8,9-tetrahydro-, (3aS,4R,10aS)-](http://img.cochemist.com/ccimg/35600/35523-89-8_b.png)
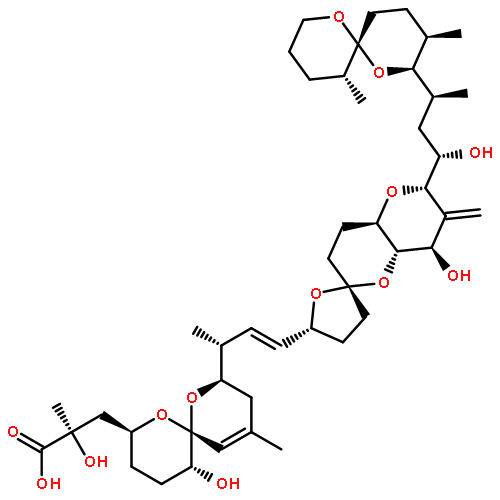
![4-Pentenoic acid,
5-[(2R,2'R,2''R,3aS,6S,6''S,7aS)-2-[(S)-[(2R,3R,5S,6S)-6-[1-[[(2R,3'R,
3aS,5S,5'S,7R,9R,9aS)-hexahydro-3',5',7-trimethylspiro[5,9-epoxy-2H-
furo[3,2-b]oxocin-2,2'-piperidin]-5(3H)-yl]methyl]ethenyl]tetrahydro-2-hy
droxy-3,5-dimethyl-2H-pyran-2-yl]hydroxymethyl]-2,3,3',3a,4',5'',6,6'',7,7
a-decahydro-4'',6-dimethyldispiro[5H-furo[3,2-b]pyran-5,2'(5'H)-furan-5'
,2''-[2H]pyran]-6''-yl]-, (4E)-](http://img.cochemist.com/ccimg/266000/265996-92-7.png)
![4-Pentenoic acid,
5-[(2R,2'R,2''R,3aS,6S,6''S,7aS)-2-[(S)-[(2R,3R,5S,6S)-6-[1-[[(2R,3'R,
3aS,5S,5'S,7R,9R,9aS)-hexahydro-3',5',7-trimethylspiro[5,9-epoxy-2H-
furo[3,2-b]oxocin-2,2'-piperidin]-5(3H)-yl]methyl]ethenyl]tetrahydro-2-hy
droxy-3,5-dimethyl-2H-pyran-2-yl]hydroxymethyl]-2,3,3',3a,4',5'',6,6'',7,7
a-decahydro-4'',6-dimethyldispiro[5H-furo[3,2-b]pyran-5,2'(5'H)-furan-5'
,2''-[2H]pyran]-6''-yl]-, (4E)-](http://img.cochemist.com/ccimg/266000/265996-92-7_b.png)