Co-reporter:Micheal L. Tuntland, L.W.-M. Fung
Analytical Biochemistry 2016 510() pp: 18-20
Publication Date(Web):1 October 2016
DOI:10.1016/j.ab.2016.07.016
Co-reporter:Anna Kim, Nina M. Wolf, Tian Zhu, Michael E. Johnson, Jiangping Deng, James L. Cook, Leslie W.-M. Fung
Bioorganic & Medicinal Chemistry 2015 23(7) pp: 1492-1499
Publication Date(Web):
DOI:10.1016/j.bmc.2015.02.016
Co-reporter:Akin Sevinc;Marta A. Witek;Leslie W. -M. Fung
Cellular & Molecular Biology Letters 2011 Volume 16( Issue 3) pp:452-461
Publication Date(Web):2011 September
DOI:10.2478/s11658-011-0017-9
Yeast two-hybrid (Y2H) and isothermal titration calorimetry (ITC) methods were used to further study the mutational effect of non-erythroid alpha spectrin (αII) at position 22 in tetramer formation with beta spectrin (βII). Four mutants, αII-V22D, V22F, V22M and V22W, were studied. For the Y2H system, we used plasmids pGBKT7, consisting of the cDNA of the first 359 residues at the N-terminal region of αII, and pGADT7, consisting of the cDNA of residues 1697–2145 at the C-terminal region of βII. Strain AH109 yeast cells were used for colony growth assays and strain Y187 was used for β-galactosidase activity assays. Y2H results showed that the C-terminal region of βII interacts with the N-terminal region of αII, either the wild type, or those with V22F, V22M or V22W mutations. The V22D mutant did not interact with βII. For ITC studies, we used recombinant proteins of the αII N-terminal fragment and of the erythroid beta spectrin (βI) C-terminal fragment; results showed that the Kd values for V22F were similar to those for the wild-type (about 7 nM), whereas the Kd values were about 35 nM for V22M and about 90 nM for V22W. We were not able to detect any binding for V22D with ITC methods. This study clearly demonstrates that the single mutation at position 22 of αII, a region critical to the function of nonerythroid α spectrin, may lead to a reduced level of spectrin tetramers and abnormal spectrin-based membrane skeleton. These abnormalities could cause abnormal neural activities in cells.
Co-reporter:Akin Sevinc;Leslie W. -M. Fung
Cellular & Molecular Biology Letters 2011 Volume 16( Issue 4) pp:
Publication Date(Web):2011 December
DOI:10.2478/s11658-011-0025-9
With yeast two-hybrid methods, we used a C-terminal fragment (residues 1697–2145) of non-erythroid beta spectrin (βII-C), including the region involved in the association with alpha spectrin to form tetramers, as the bait to screen a human brain cDNA library to identify proteins interacting with βII-C. We applied stringent selection steps to eliminate false positives and identified 17 proteins that interacted with βII-C (IPβII-C s). The proteins include a fragment (residues 38–284) of “THAP domain containing, apoptosis associated protein 3, isoform CRA g”, “glioma tumor suppressor candidate region gene 2” (residues 1-478), a fragment (residues 74–442) of septin 8 isoform c, a fragment (residues 704–953) of “coatomer protein complex, subunit beta 1, a fragment (residues 146–614) of zinc-finger protein 251, and a fragment (residues 284–435) of syntaxin binding protein 1. We used yeast three-hybrid system to determine the effects of these βII-C interacting proteins as well as of 7 proteins previously identified to interact with the tetramerization region of non-erythroid alpha spectrin (IPαII-N s) [1] on spectrin tetramer formation. The results showed that 3 IPβII-C s were able to bind βII-C even in the presence of αII-N, and 4 IPαII-N s were able to bind αII-N in the presence of βII-C. We also found that the syntaxin binding protein 1 fragment abolished αII-N and βII-C interaction, suggesting that this protein may inhibit or regulate non-erythroid spectrin tetramer formation.
Co-reporter:Ramunas Rolius;Chloe Antoniou;Lidia A. Nazarova
Cellular & Molecular Biology Letters 2010 Volume 15( Issue 3) pp:395-405
Publication Date(Web):2010 September
DOI:10.2478/s11658-010-0015-3
Calpains and caspases are ubiquitous cysteine proteases that are associated with a variety of cellular pathways. Calpains are involved in processes such as long term potentiation, cell motility and apoptosis, and have been shown to cleave non-erythroid (brain) α- and β-spectrin and erythroid β-spectrin. The cleavage of erythroid α-spectrin by calpain has not been reported. Caspases play an important role in the initiation and execution of apoptosis, and have been shown to cleave non-erythroid but not erythroid spectrin. We have studied the effect of spectrin fragments on calpain and caspase activities. The erythroid and non-erythroid spectrin fragments used were from the N-terminal region of α-spectrin, and C-terminal region of β-spectrin, both consisting of regions involved in spectrin tetramer formation. We observed that the all spectrin fragments exhibited a concentration-dependent inhibitory effect on calpain, but not caspase activity. It is clear that additional studies are warranted to determine the physiological significance of calpain inhibition by spectrin fragments. Our findings suggest that calpain activity is modulated by the presence of spectrin partial domains at the tetramerization site. It is not clear whether the inhibitory effect is substrate specific or is a general effect. Further studies of this inhibitory effect may lead to the identification and development of new therapeutic agents specifically for calpains, but not for caspases. Proteins/peptides with a coiled coil helical conformation should be studied for potential inhibitory effects on calpain activity.
Co-reporter:Jianxia Kang;Yuanli Song;Akin Sevinc
Cellular & Molecular Biology Letters 2010 Volume 15( Issue 1) pp:
Publication Date(Web):2010 March
DOI:10.2478/s11658-009-0031-3
Spectrin tetramerization is important for the erythrocyte to maintain its unique shape, elasticity and deformability. We used recombinant model proteins to show the importance of one residue (G46) in the erythroid α-spectrin junction region that affects spectrin tetramer formation. The G46 residue in the erythroid spectrin N-terminal junction region is the only residue that differs from that in non-erythroid spectrin. The corresponding residue is R37. We believe that this difference may be, at least in part, responsible for the 15-fold difference in the equilibrium constants of erythroid and non-erythroid tetramer formation. In this study, we replaced the Gly residue with Ala, Arg or Glu residues in an erythroid α-spectrin model protein to give G46A, G46R or G46E, respectively. We found that their association affinities with a β-spectrin model protein were quite different from each other. G46R exhibited a 10-fold increase and G46E exhibited a 16-fold decrease, whereas G46A showed little difference, when compared with the wild type. The thermal and urea denaturation experiments showed insignificant structural change in G46R. Thus, the differences in affinity were due to differences in local, specific interactions, rather than conformational differences in these variants. An intra-helical salt bridge in G46R may stabilize the partial domain single helix in α-spectrin, Helix C’, to allow a more stable helical bundling in the αβ complex in spectrin tetramers. These results not only showed the importance of residue G46 in erythroid α-spectrin, but also provided insights toward the differences in association affinity between erythroid and non-erythroid spectrin to form spectrin tetramers.
Co-reporter:Qufei Li and L. W.-M. Fung
Biochemistry 2009 Volume 48(Issue 1) pp:
Publication Date(Web):December 12, 2008
DOI:10.1021/bi8013032
The N-terminal region of α-spectrin is responsible for its association with β-spectrin in a heterodimer, forming functional tetramers. Non-erythroid α-spectrin (αII-spectrin) has a significantly higher association affinity for β-spectrin than the homologous erythroid α-spectrin (αI-spectrin). We have previously determined the solution structure of the N-terminal region of αI-spectrin by NMR methods, but currently no structural information is available for αII-spectrin. We have used cysteine scanning, spin labeling electron paramagnetic resonance (EPR), and isothermal titration calorimetry (ITC) methods to study the tetramerization region of αII-spectrin. EPR data clearly show that, in αII-spectrin, the first nine N-terminal residues were unstructured, followed by an irregular helix (helix C′), frayed at the N-terminal end, but rigid at the C-terminal end, which merges into the putative triple-helical structural domain. The region corresponding to the important unstructured junction region linking helix C′ to the first structural domain in αI-spectrin was clearly structured. On the basis of the published model for aligning helices A′, B′, and C′, important interactions among residues in helix C′ of αI- and αII-spectrin and helices A′ and B′ of βI- and βII-spectrin are identified, suggesting similar coiled coil helical bundling for spectrin I and II in forming tetramers. The differences in affinity are likely due to the differences in the conformation of the junction regions. Equilibrium dissociation constants of spin-labeled αII and βI complexes from ITC measurements indicate that residues 15, 19, 37, and 40 are functionally important residues in αII-spectrin. Interestingly, all four corresponding homologous residues in αI-spectrin (residues 24, 28, 46, and 49) have been reported to be clinically significant residues involved in hematological diseases.
Co-reporter:Chloe Antoniou, Vinh Q. Lam and L. W.-M. Fung
Biochemistry 2008 Volume 47(Issue 40) pp:
Publication Date(Web):September 11, 2008
DOI:10.1021/bi800840p
We used cysteine scanning, isothermal titration calorimetry (ITC) and spin label EPR methods to study the two regions that flank the partial domain Helix C′ of the N-terminal end of α-spectrin (residues 14−20 and residues 44−54) in the absence and presence of a model protein of the β-spectrin C-terminal end. In the absence of β-spectrin, residues 14−20 and 46−52 were known to be unstructured. The EPR spectral values of the inverse line width (ΔH−1) and of the width between the low field peak and the central peak (aZ) of residues in part of the first unstructured region (residues 17−20) and of most residues in the second unstructured junction region (residues 46−52) changed dramatically upon association with β-spectrin, suggesting that the two regions undergo a conformational change, becoming more rigid and likely becoming helical. ITC results showed that three of the seven residues in the junction region (residues 46−52) were very important in its association with β-spectrin, in the following order: L49 > G46 > K48. In general, our results suggest that any mutations that affect the propensity of helical formation in the region spanning residues 17−52 in α-spectrin, or that affect hydrophobic clustering and/or salt-bridge stabilization of the bundled helices, would affect spectrin tetramer formation, and may lead to blood disorders.
Co-reporter:Chloe Antoniou, L.W.-M. Fung
Analytical Biochemistry 2008 Volume 376(Issue 1) pp:160-162
Publication Date(Web):1 May 2008
DOI:10.1016/j.ab.2008.02.001
Site-directed spin labeling electron paramagnetic resonance methods have been an important tool in studying protein–protein interactions. Labels are often attached to a cysteine residue, and spectra are acquired with and without binding partner(s) to provide information on the binding. This requires a knowledge of the label location which is simplified if the label remains faithfully attached to the designated residue in the complex. We report a system where this is not the case because the label was extracted by dialysis-resistant glutathione molecules. Once this artifact is identified, spectral subtraction provides a solution for meaningful data interpretation.
Co-reporter:Younsang Oh;Leslie W. -M. Fung
Cellular & Molecular Biology Letters 2007 Volume 12( Issue 4) pp:604-620
Publication Date(Web):2007 December
DOI:10.2478/s11658-007-0028-8
The N-terminal region of non-erythroid alpha spectrin (SpαII) is responsible for interacting with its binding partner, beta spectrin, to form functional spectrin tetramers. We used a yeast-two-hybrid system, with an N-terminal segment of alpha spectrin representing the functional tetramerization site, as a bait to screen human brain c-DNA library for proteins that interact with the alpha spectrin segment. In addition to several beta spectrin isoforms, we identified 14 proteins that interact with SpαII. Seven of the 14 were matched to 6 known proteins: Duo protein, Lysyl-tRNA synthetase, TBP associated factor 1, two isoforms (b and c) of a protein kinase A interacting protein and Zinc finger protein 333 (2 different segments). Four of the 6 proteins are located primarily in the nucleus, suggesting that spectrin plays important roles in nuclear functions. The remaining 7 proteins were unknown to the protein data base. Structural predictions show that many of the 14 proteins consist of a large portion of unstructured regions, suggesting that many of these proteins fold into a rather flexible conformation. It is interesting to note that all but 3 of the 14 proteins are predicted to consist of one to four coiled coils (amphiphilic helices). A mutation in SpαII, V22D, which interferes with the coiled coil bundling of SpαII with beta spectrin, also affects SpαII interaction with Duo protein, TBP associated factor 1 and Lysyl-tRNA synthetase, suggesting that they may compete with beta spectrin for interaction with SpαII. Future structural and functional studies of these proteins to provide interaction mechanisms will no doubt lead to a better understanding of brain physiology and pathophysiology.
Co-reporter:Marta A. Witek, L.W.-M. Fung
Brain Research (2 October 2013) Volume 1533() pp:1-15
Publication Date(Web):2 October 2013
DOI:10.1016/j.brainres.2013.08.010
•Spectrin breakdown products are detected under various brain conditions.•Kinetics studies of breakdown reactions of spectrin model proteins by caspase-3.•More efficient cleavage to generate the 150 kDa fragment than the 120 kDa fragment.•The formations of 150 kDa and 120 kDa fragments are independent of each other.•Development of more precise biomarkers to monitor spectrin degradation.Under various physiological and patho-physiological conditions, spectrin breakdown reactions generate several spectrin breakdown products (SBDPs)—in particular SBDPs of 150 kDa (SBDP150) and 120 kDa (SBDP120). Recently, numerous studies have shown that reactions leading to SBDPs are physiologically relevant, well regulated, and complex. Yet molecular studies on the mechanism of the SBDP formation are comparatively scarce. We have designed basic systems to allow us to follow the breakdown of αII-spectrin model proteins by caspase-3 in detail with gel electrophoresis, fluorescence and mass spectrometry methods. Amongst the predicted and reported sites, our results show that caspase-3 cleaves after residues D1185 and D1478, but not after residues D888, D1340 and D1475. We also found that the cleavage at these two sites is independent of each other. It may be possible to inhibit one site without affecting the other site. Cleavage after residue D1185 in intact αII-spectrin leads to SBDP150, and cleavage after D1478 site leads to SBDP120. Our results also show that the cleavage after the D1185 residue is unusually efficient, with a kcat/KM value of 40,000 M−1 s−1, and the cleavage after the D1478 site is more similar to most of the other reported caspase-3 substrates, with a kcat/KM value of 3000 M−1 s−1. We believe that this study lays out a methodology and foundation to study caspase-3 catalyzed spectrin breakdown to provide quantitative information. Molecular understanding may lead to better understanding of brain injuries and more precise and specific biomarker development.

![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8.png)
![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8_b.png)
![1-[5-(2-CHLOROPHENYL)-1,2-OXAZOL-3-YL]METHANAMINE HYDROCHLORIDE (1:1)](http://img.cochemist.com/ccimg/1160300/1160245-59-9.png)
![1-[5-(2-CHLOROPHENYL)-1,2-OXAZOL-3-YL]METHANAMINE HYDROCHLORIDE (1:1)](http://img.cochemist.com/ccimg/1160300/1160245-59-9_b.png)






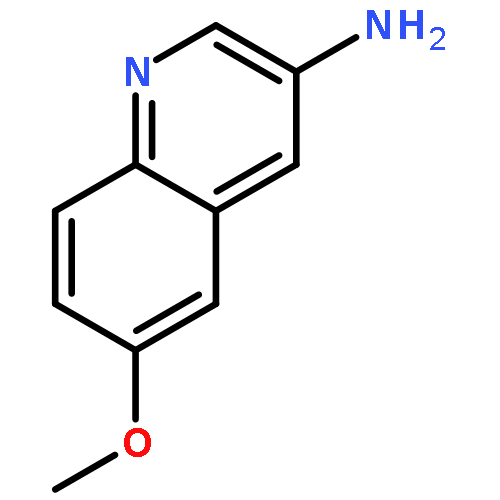
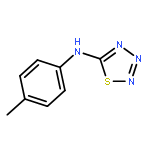
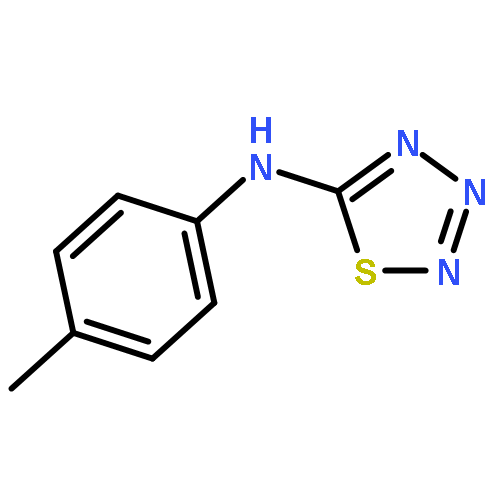


![3-Phenyl-[1,2,4]triazolo[3,4-b][1,3,4]thiadiazol-6-amine](http://img.cochemist.com/ccimg/3200/3176-53-2.png)
![3-Phenyl-[1,2,4]triazolo[3,4-b][1,3,4]thiadiazol-6-amine](http://img.cochemist.com/ccimg/3200/3176-53-2_b.png)
![L-Aspartic acid,N-[[5-amino-1-(5-O-phosphono-b-D-ribofuranosyl)-1H-imidazol-4-yl]carbonyl]-](http://img.cochemist.com/ccimg/3100/3031-95-6.png)
![L-Aspartic acid,N-[[5-amino-1-(5-O-phosphono-b-D-ribofuranosyl)-1H-imidazol-4-yl]carbonyl]-](http://img.cochemist.com/ccimg/3100/3031-95-6_b.png)