Co-reporter:Sajjad Hossain and Elizabeth M. Boon
ACS Infectious Diseases June 9, 2017 Volume 3(Issue 6) pp:454-454
Publication Date(Web):February 27, 2017
DOI:10.1021/acsinfecdis.7b00027
Nitric oxide (NO) is a radical diatomic gas molecule that, at low concentrations, plays important signaling roles in both eukaryotes and bacteria. In recent years, it has become evident that bacteria respond to low levels of NO in order to modulate their group behavior. Many bacteria respond via NO ligation to a well-established NO sensor called H-NOX (heme-nitric oxide/oxygen binding domain). Many others, such as Pseudomonas aeruginosa, lack an annotated hnoX gene in their genome yet are able to respond to low levels of NO to disperse their biofilms. This suggests the existence of a previously uncharacterized NO sensor. In this study, we describe the discovery of a novel nitric oxide binding protein (NosP; NO-sensing protein), which is much more widely conserved in bacteria than H-NOX, as well as a novel NO-responsive pathway in P. aeruginosa. We demonstrate that biofilms of a P. aeruginosa mutant lacking components of the NosP pathway lose the ability to disperse in response to NO. Upon cloning, expressing, and purifying NosP, we find it binds heme and ligates to NO with a dissociation rate constant that is comparable to that of other well-established NO-sensing proteins. Moreover, we show that NO-bound NosP is able to regulate the phosphorelay activity of a hybrid histidine kinase that is involved in biofilm regulation in P. aeruginosa. Thus, here, we present evidence of a novel NO-responsive pathway that regulates biofilm in P. aeruginosa.Keywords: biofilm; nitric oxide; nitric oxide sensor; nitric oxide signaling; nitric oxide-responsive kinase; NosP;
Co-reporter:Sajjad Hossain, Lisa-Marie Nisbett, and Elizabeth M. Boon
Accounts of Chemical Research July 18, 2017 Volume 50(Issue 7) pp:1633-1633
Publication Date(Web):June 12, 2017
DOI:10.1021/acs.accounts.7b00095
ConspectusBacterial biofilms form when bacteria adhere to a surface and produce an exopolysaccharide matrix (Costerton Science 1999, 284, 1318; Davies Science 1998, 280, 295; Flemming Nat. Rev. Microbiol. 2010, 8, 623). Because biofilms are resistant to antibiotics, they are problematic in many aspects of human health and welfare, causing, for instance, persistent fouling of medical implants such as catheters and artificial joints (Brunetto Chimia 2008, 62, 249). They are responsible for chronic infections in the lungs of cystic fibrosis patients and in open wounds, such as those associated with burns and diabetes. They are also a major contributor to hospital-acquired infections (Sievert Infec. Control Hosp. Epidemiol. 2013, 34, 1; Tatterson Front. Biosci. 2001, 6, D890).It has been hypothesized that effective methods of biofilm control will have widespread application (Landini Appl. Microbiol. Biotechnol. 2010, 86, 813). A promising strategy is to target the mechanisms that drive biofilm dispersal, because dispersal results in biofilm removal and in the restoration of antibiotic sensitivity. First documented in Nitrosomonas europaea (Schmidt J. Bacteriol. 2004, 186, 2781) and the cystic fibrosis-associated pathogen Pseudomonas aeruginosa (Barraud J. Bacteriol. 2006, 188, 7344; J. Bacteriol. 2009, 191, 7333), regulation of biofilm formation by nanomolar levels of the diatomic gas nitric oxide (NO) has now been documented in numerous bacteria (Barraud Microb. Biotechnol. 2009, 2, 370; McDougald Nat. Rev. Microbiol. 2012, 10, 39; Arora Biochemistry 2015, 54, 3717; Barraud Curr. Pharm. Des. 2015, 21, 31). NO-mediated pathways are, therefore, promising candidates for biofilm regulation. Characterization of the NO sensors and NO-regulated signaling pathways should allow for rational manipulation of these pathways for therapeutic applications.Several laboratories, including our own, have shown that a class of NO sensors called H-NOX (heme-nitric oxide or oxygen binding domain) affects biofilm formation by regulating intracellular cyclic di-GMP concentrations and quorum sensing (Arora Biochemistry 2015, 54, 3717; Plate Trends Biochem. Sci. 2013, 38, 566; Nisbett Biochemistry 2016, 55, 4873). Many bacteria that respond to NO do not encode an hnoX gene, however. My laboratory has now discovered an additional family of bacterial NO sensors, called NosP (nitric oxide sensing protein). Importantly, NosP domains are widely conserved in bacteria, especially Gram-negative bacteria, where they are encoded as fusions with or in close chromosomal proximity to histidine kinases or cyclic di-GMP synthesis or phosphodiesterase enzyme, consistent with signaling. In this Account, we briefly review NO and H-NOX signaling in bacterial biofilms, describe our discovery of the NosP family, and provide support for its role in biofilm regulation in Pseudomonas aeruginosa, Vibrio cholerae, Legionella pneumophila, and Shewanella oneidensis.
Co-reporter:Lisa-Marie Nisbett and Elizabeth M. Boon
Biochemistry 2016 Volume 55(Issue 35) pp:4873
Publication Date(Web):August 1, 2016
DOI:10.1021/acs.biochem.6b00754
Nitric oxide (NO) is a freely diffusible, radical gas that has now been established as an integral signaling molecule in eukaryotes and bacteria. It has been demonstrated that NO signaling is initiated upon ligation to the heme iron of an H-NOX domain in mammals and in some bacteria. Bacterial H-NOX proteins have been found to interact with enzymes that participate in signaling pathways and regulate bacterial processes such as quorum sensing, biofilm formation, and symbiosis. Here, we review the biochemical characterization of these signaling pathways and, where available, describe how ligation of NO to H-NOX specifically regulates the activity of these pathways and their associated bacterial phenotypes.
Co-reporter:Dhruv P. Arora, Sajjad Hossain, Yueming Xu, and Elizabeth M. Boon
Biochemistry 2015 Volume 54(Issue 24) pp:3717-3728
Publication Date(Web):May 21, 2015
DOI:10.1021/bi501476n
Biofilms are surface-associated, multicellular communities of bacteria. Once established, they are extremely difficult to eradicate by antimicrobial treatment. It has been demonstrated in many species that biofilm formation may be regulated by the diatomic signaling molecule nitric oxide (NO). Although this is still a relatively new area of research, we review here the literature reporting an effect of NO on bacterial biofilm formation, emphasizing dose-dependent responses to NO concentrations when possible. Where it has been investigated, the underlying NO sensors or signaling pathways are also discussed. Most of the examples of NO-mediated biofilm regulation have been documented with exogenously applied NO, but we also survey possible natural sources of NO in biofilm regulation, including endogenously generated NO. Finally, because of the apparent broad-spectrum, antibiofilm effects of NO, NO-releasing materials and prodrugs have also been explored in this minireview.
Co-reporter:Tanaya Lahiri, Bowu Luan, Daniel P. Raleigh, and Elizabeth M. Boon
Biochemistry 2014 Volume 53(Issue 13) pp:
Publication Date(Web):March 15, 2014
DOI:10.1021/bi401597m
Biofilms are surface-attached communities of bacteria enclosed in a polysaccharide matrix. Bacteria in a biofilm are extremely resistant to antibiotics. Several recent reports have linked the signaling molecule nitric oxide (NO) with biofilm dispersal. We have previously reported that an H-NOX (heme-nitric oxide/oxygen binding) protein in the biofilm-dwelling bacterium Shewanella woodyi mediates NO-induced biofilm dispersal. In S. woodyi, H-NOX (SwH-NOX) is cocistronic with a gene encoding a dual-functioning diguanylate cyclase/phosphodiesterase enzyme, designated here as HaCE (H-NOX-associated cyclic-di-GMP processing enzyme). Enzymes such as these are responsible for regulating the intracellular concentrations of cyclic-di-GMP, a secondary signaling molecule essential to biofilm formation in bacteria. We have demonstrated that NO-bound SwH-NOX regulates both enzymatic activities of SwHaCE, resulting in decreased cellular cyclic-di-GMP levels and disruption of biofilm formation. Thus, H-NOX/HaCE represents a potential drug target for regulating biofilm formation. In this work, the SwH-NOX surface residues critical for the formation of a protein complex with SwHaCE are identified using nuclear magnetic resonance, fluorescence quenching, and cosedimentation. Enzyme assays confirm this protein–protein interface and its importance for H-NOX/HaCE function.
Co-reporter:John G. Kosowicz, Elizabeth M. Boon
Journal of Inorganic Biochemistry 2013 Volume 126() pp:91-95
Publication Date(Web):September 2013
DOI:10.1016/j.jinorgbio.2013.05.012
•Fluoride is used to probe distal pocket H-bonding in the H-NOX family.•The role of each member of the distal pocket H-bonding network is characterized.•Distal pocket H-bonding regulates ligand binding in H-NOX.•Distal pocket molecular crowding plays a role in fine-tuning ligand binding to H-NOX.•Supports the hypothesis that H-bonding underlies sGC ligand discrimination.Nitric oxide (NO) and dioxygen (O2) are gases of similar size, shape, and electrostatic potential, but different physiological function. In aerobic organisms, the cellular concentration of O2 far exceeds that of NO; instead NO relies heavily on the ability of its receptor to discriminate against O2. In mammals, soluble guanylate cyclase (sGC) serves this role, binding NO with picomolar sensitivity and excluding O2 binding. Interestingly, some bacterial homologs of sGC, including the H-NOX (heme-nitric oxide/oxygen) domain from Thermoanaerobacter tengcongensis, tightly bind O2. Three distal pocket residues (Trp9, Asn74, and Tyr140) form a hydrogen-bonding network that stabilizes O2 binding to TtH-NOX. Therefore, a current hypothesis to explain sGC ligand specificity is that sGC lacks H-bond donors that preferentially stabilize O2 binding. The wavelength maximum of the charge-transfer band (CT1) in the electronic spectrum of the fluoride complex of ferric hemoproteins is a sensitive probe of H-bonding. Here, in order to gain further understanding of the distal pocket H-bonding network in TtH-NOX, we employ fluoride as a spectroscopic probe. As expected, our results indicate that Y140 donates a strong H-bond to the heme-bound ligand. We find that an H-bond from Asn74 as well as distal pocket crowding contributes to positioning Tyr140 for a strong and directed H-bond to iron-bound ligands; indeed crowding may be the primary role for Trp9. We clarify the role of H-bonding in sGC ligand discrimination and suggest that sterics also regulate ligand binding in the H-NOX family.Analysis of the UV/visible spectra of ferric-fluoride complexes of wild-type and mutant H-NOX (heme-nitric oxide/oxygen binding) proteins yields an understanding of the role of the distal pocket H-bonding network in regulating ligand binding.
Co-reporter:Sandhya Muralidharan
Journal of the American Chemical Society 2012 Volume 134(Issue 4) pp:2044-2046
Publication Date(Web):January 17, 2012
DOI:10.1021/ja211576b
The H-NOX family of nitric oxide (NO) sensing proteins has received considerable attention because its members include the mammalian NO sensor, soluble guanylate cyclase. Despite this attention, the mechanism of signal transduction has not been elucidated. Structural studies of bacterial members of the family have revealed that the H-NOX heme cofactor is extremely distorted from planarity. Furthermore, it has been determined that heme distortion is maintained primarily by a conserved proline residue located in the proximal heme pocket. It has been suggested that changes in heme planarity may contribute to signal transduction. Here we demonstrate that heme flattening is, indeed, sufficient for signal transduction in the H-NOX family. Using our previously described H-NOX/diguanylate cyclase functional partners from Shewanella woodyi, we demonstrate that mutation of the conserved proline (P117 in SwH-NOX) to alanine, which results in heme flattening, has the same affect on phosphodiesterase activity as NO binding to wildtype SwH-NOX. This study demonstrates, for the first time, that heme flattening mimics the activated, NO-bound state of H-NOX and suggests that NO binding induces heme flattening as part of the signal transduction mechanism in the H-NOX family.
Co-reporter:Bernadette M. Henares, Kate E. Higgins, and Elizabeth M. Boon
ACS Chemical Biology 2012 Volume 7(Issue 8) pp:1331
Publication Date(Web):May 19, 2012
DOI:10.1021/cb300215t
Bacteria use small molecules to assess the density and identity of nearby organisms and formulate a response. This process, called quorum sensing (QS), commonly regulates bioluminescence, biofilm formation, and virulence. Vibrio harveyi have three described QS circuits. Each involves the synthesis of a molecule that regulates phosphorylation of its cognate receptor kinase. Each receptor exchanges phosphate with a common phosphorelay protein, LuxU, which ultimately regulates bioluminescence. Here, we show that another small molecule, nitric oxide (NO), participates in QS through LuxU. V. harveyi display a NO concentration-dependent increase in bioluminescence that is regulated by an hnoX gene. We demonstrate that H-NOX is a NO sensor and NO/H-NOX regulates phosphorylation of a kinase that transfers phosphate to LuxU. This study reveals the discovery of a fourth QS pathway in V. harveyi and suggests that bacteria use QS to integrate not only the density of bacteria but also other diverse information about their environment into decisions about gene expression.
Co-reporter:Zhou Dai, Erik R. Farquhar, Dhruv P. Arora and Elizabeth M. Boon
Dalton Transactions 2012 vol. 41(Issue 26) pp:7984-7993
Publication Date(Web):17 Feb 2012
DOI:10.1039/C2DT30147D
The H-NOX (Heme–Nitric oxide/OXygen binding) family of diatomic gas sensing hemoproteins has attracted great interest. Soluble guanylate cyclase (sGC), the well-characterized eukaryotic nitric oxide (NO) sensor is an H-NOX family member. When NO binds sGC at the ferrous histidine-ligated protoporphyrin-IX, the proximal histidine ligand dissociates, resulting in a 5-coordinate (5c) complex; formation of this 5c complex is viewed as necessary for activation of sGC. Characterization of other H-NOX family members has revealed that while most also bind NO in a 5c complex, some bind NO in a 6-coordinate (6c) complex or as a 5c/6c mixture. To gain insight into the heme pocket structural differences between 5c and 6c Fe(II)–NO H-NOX complexes, we investigated the extended X-ray absorption fine structure (EXAFS) of the Fe(II)–unligated and Fe(II)–NO complexes of H-NOX domains from three species, Thermoanaerobacter tengcongensis, Shewanella woodyi, and Pseudoalteromonas atlantica. Although the Fe(II)–NO complex of TtH-NOX is formally 6c, we found the Fe–NHis bond is substantially lengthened. Furthermore, although NO binds to SwH-NOX and PaH-NOX as a 5c complex, consistent with histidine dissociation, the EXAFS data do not exclude a very weakly associated histidine. Regardless of coordination number, upon NO-binding, the Fe–Nporphyrin bond lengths in all three H-NOXs contract by ∼0.07 Å. This study reveals that the overall heme structure of 5c and 6c Fe(II)–NO H-NOX complexes are substantially similar, suggesting that formal histidine dissociation may not be required to trigger NO/H-NOX signal transduction. The study has refined our understanding of the molecular mechanisms underlying NO/H-NOX signaling.
Co-reporter:Niu Liu, Yueming Xu, Sajjad Hossain, Nick Huang, Dan Coursolle, Jeffrey A. Gralnick, and Elizabeth M. Boon
Biochemistry 2012 Volume 51(Issue 10) pp:
Publication Date(Web):February 23, 2012
DOI:10.1021/bi201753f
Although several reports have documented nitric oxide (NO) regulation of biofilm formation, the molecular basis of this phenomenon is unknown. In many bacteria, an H-NOX (heme-nitric oxide/oxygen-binding) gene is found near a diguanylate cyclase (DGC) gene. H-NOX domains are conserved hemoproteins that are known NO sensors. It is widely recognized that cyclic di-GMP (c-di-GMP) is a ubiquitous bacterial signaling molecule that regulates the transition between motility and biofilm. Therefore, NO may influence biofilm formation through H-NOX regulation of DGC, thus providing a molecular-level explanation for NO regulation of biofilm formation. This work demonstrates that, indeed, NO-bound H-NOX negatively affects biofilm formation by directly regulating c-di-GMP turnover in Shewanella woodyi strain MS32. Exposure of wild-type S. woodyi to a nanomolar level of NO resulted in the formation of thinner biofilms, and less intracellular c-di-GMP, than in the absence of NO. Also, a mutant strain in the gene encoding SwH-NOX showed a decreased level of biofilm formation (and a decreased amount of intracellular c-di-GMP) with no change observed upon NO addition. Furthermore, using purified proteins, it was demonstrated that SwH-NOX and SwDGC are binding partners. SwDGC is a dual-functioning DGC; it has diguanylate cyclase and phosphodiesterase activities. These data indicate that NO-bound SwH-NOX enhances c-di-GMP degradation, but not synthesis, by SwDGC. These results support the biofilm growth data and indicate that S. woodyi senses nanomolar NO with an H-NOX domain and that SwH-NOX regulates SwDGC activity, resulting in a reduction in c-di-GMP concentration and a decreased level of biofilm growth in the presence of NO. These data provide a detailed molecular mechanism for NO regulation of c-di-GMP signaling and biofilm formation.
Co-reporter:Zhou Dai, Elizabeth M. Boon
Journal of Inorganic Biochemistry 2011 Volume 105(Issue 6) pp:784-792
Publication Date(Web):June 2011
DOI:10.1016/j.jinorgbio.2011.03.002
Heme-Nitric oxide and/or OXygen binding (H-NOX) proteins are a family of diatomic gas binding hemoproteins that have attracted intense research interest. Here we employ X-ray absorption near-edge structure (XANES) spectroscopy to study the nitric oxide (NO) binding site of H-NOX. This is the first time this technique has been utilized to examine the NO/H-NOX signaling pathway. XANES spectra of wildtype and a point mutant (proline 115 to alanine, P115A) of the H-NOX domain from Thermoanaerobacter tengcongensis (Tt H-NOX) were obtained and analyzed for ferrous and ferric complexes of the protein. This work provides specific structural characterization of the solution state of several Tt H-NOX ferrous complexes (− unligated, − NO, and − CO) that were previously unavailable. Our iron K-edges indicate effective charge on the iron center in the various complexes and report on the electronic environment of heme iron. We analyzed the ligand field indicator ratio (LFIR), which is extracted from XANES spectra, for each complex, providing an understanding of ligand field strength, spin state of the central iron, movement of the iron atom upon ligation, and ligand binding properties. In particular, our LFIRs indicate that the heme iron is dramatically displaced towards the distal pocket during ligand binding. Based on these results, we propose that iron displacement towards the distal heme pocket is an essential step in signal initiation in H-NOX proteins. This provides a mechanistic link between ligand binding and the changes in heme and protein conformation that have been observed for H-NOX family members during signaling.A XANES analysis of TtH-NOX complexes reveals that upon ligand binding, iron moves into the distal pocket, thus initiating signaling.
Co-reporter:Zhou Dai
Journal of the American Chemical Society 2010 Volume 132(Issue 33) pp:11496-11503
Publication Date(Web):August 4, 2010
DOI:10.1021/ja101674z
A new cyanide sensing system, the Heme-Nitric oxide and/or OXygen binding domain (H-NOX domain) from Thermoanaerobacter tengcongensis (Tt H-NOX), has been investigated. With straightforward absorbance-based detection, we have achieved a cyanide detection limit of 0.5 μM (∼10 ppb) with an upper detection range that is adjustable with protein concentration. We find a linear correlation of multiple spectroscopic features with cyanide concentration. These spectroscopic features include the Soret band maximum and absorbance changes in both the Soret and α/β band regions of the spectrum. Multiple probes for cyanide detection makes sensing with Tt H-NOX unique compared to other cyanide sensing methods. Furthermore, using site-directed mutagenesis, we have rationally engineered the heme pocket of Tt H-NOX to improve its cyanide sensing properties. Using a mutant that alters the heme structure of Tt H-NOX (P115A) we were able to introduce colorimetric detection of cyanide. Through substituting phenylalanine 78 with a smaller (valine, F78V) or a larger residue (tyrosine, F78Y), we demonstrate a correlation with distal pocket steric crowding and affinity for cyanide. In particular, F78V Tt H-NOX shows a significant increase in CN− binding affinity and selectivity. Thus, we demonstrate the ability to fine-tune the affinity and specificity of Tt H-NOX for cyanide, suggesting that Tt H-NOX can be readily tailored into a practical cyanide sensor.
Co-reporter:Niu Liu, Taemee Pak and Elizabeth M. Boon
Molecular BioSystems 2010 vol. 6(Issue 9) pp:1561-1564
Publication Date(Web):13 May 2010
DOI:10.1039/C002246B
Cyclic-di-GMP signaling is used by many bacteria to control biofilm formation. As biofilm formation is an important survival strategy for many bacteria, the synthesis and degradation of cyclic-di-GMP is tightly regulated by enzymes containing domains with conserved GGDEF and EAL sequence motifs, respectively. In this report we characterize a protein with both cyclase and phosphodiesterase activities and demonstrate that it contributes to secretion of the extracellular polysaccharide matrix, an important step in early biofilm formation.
Co-reporter:Natasha M. Nesbitt, Dhruv P. Arora, Roger A. Johnson, Elizabeth M. Boon
Biotechnology Reports (September 2015) Volume 7() pp:
Publication Date(Web):1 September 2015
DOI:10.1016/j.btre.2015.04.008
Cyclic-diGMP is a bacterial messenger that regulates many physiological processes, including many attributed to pathogenicity. Bacteria synthesize cyclic-diGMP from GTP using diguanylate cyclases; its hydrolysis is catalyzed by phosphodiesterases. Here we report the over-expression and purification of a bi-functional diguanylate cyclase-phosphodiesterase from Agrobacterium vitis S4. Using homology modeling and primary structure alignment, we identify several amino acids predicted to participate in the phosphodiesterase reaction. Upon altering selected residues, we obtain variants of the enzyme that efficiently and quantitatively catalyze the synthesis of cyclic-diGMP from GTP without hydrolysis to pGpG. Additionally, we identify a variant that produces cyclic-diGMP while immobilized to NiNTA beads and can catalyze the conversion of [α-32P]-GTP to [32P]-cyclic-diGMP. In short, we characterize a novel cyclic-diGMP processing enzyme and demonstrate its utility for efficient and cost-effective production of cyclic-diGMP, as well as modified cyclic-diGMP molecules, for use as probes in studying the many important biological processes mediated by cyclic-diGMP.
Download full-size image
Co-reporter:Zhou Dai, Erik R. Farquhar, Dhruv P. Arora and Elizabeth M. Boon
Dalton Transactions 2012 - vol. 41(Issue 26) pp:NaN7993-7993
Publication Date(Web):2012/02/17
DOI:10.1039/C2DT30147D
The H-NOX (Heme–Nitric oxide/OXygen binding) family of diatomic gas sensing hemoproteins has attracted great interest. Soluble guanylate cyclase (sGC), the well-characterized eukaryotic nitric oxide (NO) sensor is an H-NOX family member. When NO binds sGC at the ferrous histidine-ligated protoporphyrin-IX, the proximal histidine ligand dissociates, resulting in a 5-coordinate (5c) complex; formation of this 5c complex is viewed as necessary for activation of sGC. Characterization of other H-NOX family members has revealed that while most also bind NO in a 5c complex, some bind NO in a 6-coordinate (6c) complex or as a 5c/6c mixture. To gain insight into the heme pocket structural differences between 5c and 6c Fe(II)–NO H-NOX complexes, we investigated the extended X-ray absorption fine structure (EXAFS) of the Fe(II)–unligated and Fe(II)–NO complexes of H-NOX domains from three species, Thermoanaerobacter tengcongensis, Shewanella woodyi, and Pseudoalteromonas atlantica. Although the Fe(II)–NO complex of TtH-NOX is formally 6c, we found the Fe–NHis bond is substantially lengthened. Furthermore, although NO binds to SwH-NOX and PaH-NOX as a 5c complex, consistent with histidine dissociation, the EXAFS data do not exclude a very weakly associated histidine. Regardless of coordination number, upon NO-binding, the Fe–Nporphyrin bond lengths in all three H-NOXs contract by ∼0.07 Å. This study reveals that the overall heme structure of 5c and 6c Fe(II)–NO H-NOX complexes are substantially similar, suggesting that formal histidine dissociation may not be required to trigger NO/H-NOX signal transduction. The study has refined our understanding of the molecular mechanisms underlying NO/H-NOX signaling.
.jpg)



![Benzene, 1-bromo-3-[[3-(bromomethyl)phenyl]thio]-](http://img.cochemist.com/ccimg/926700/926622-38-0.png)
![Benzene, 1-bromo-3-[[3-(bromomethyl)phenyl]thio]-](http://img.cochemist.com/ccimg/926700/926622-38-0_b.png)
![Benzenemethanol, 3-[(3-bromophenyl)thio]-](http://img.cochemist.com/ccimg/926700/926622-36-8.png)
![Benzenemethanol, 3-[(3-bromophenyl)thio]-](http://img.cochemist.com/ccimg/926700/926622-36-8_b.png)


![BENZENEACETAMIDE, N-[[(2-MERCAPTOETHYL)THIO]METHYL]-](http://img.cochemist.com/ccimg/852000/851987-01-4.png)
![BENZENEACETAMIDE, N-[[(2-MERCAPTOETHYL)THIO]METHYL]-](http://img.cochemist.com/ccimg/852000/851987-01-4_b.png)




![2-morpholin-4-ylpyrimido[2,1-a]isoquinolin-4-one](http://img.cochemist.com/ccimg/168500/168425-64-7.png)
![2-morpholin-4-ylpyrimido[2,1-a]isoquinolin-4-one](http://img.cochemist.com/ccimg/168500/168425-64-7_b.png)


amino]-, ethyl ester,(3S)-](/data/chemimg/115300/152673-22-8.png)
amino]-, ethyl ester,(3S)-](/data/chemimg/115300/152673-22-8_b.png)






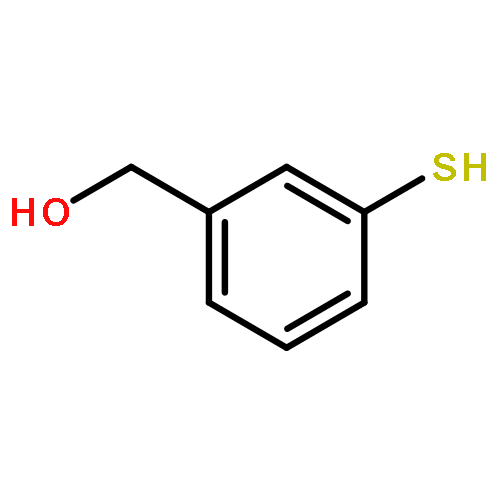
![4,5-Dioxo-4,5-dihydro-1H-pyrrolo[2,3-f]quinoline-2,7,9-tricarboxylic acid](/data/chemimg/654800/72909-34-3.png)
![4,5-Dioxo-4,5-dihydro-1H-pyrrolo[2,3-f]quinoline-2,7,9-tricarboxylic acid](/data/chemimg/654800/72909-34-3_b.png)


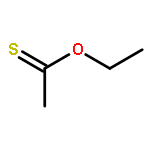

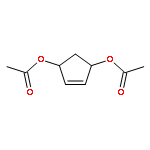

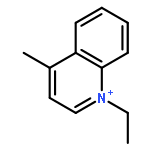

![Butanamide,2-hydroxy-N-[3-[(2-mercaptoethyl)amino]-3-oxopropyl]-3,3-dimethyl-4-(phosphonooxy)-,(2R)-](http://img.cochemist.com/ccimg/2300/2226-71-3.png)
![Butanamide,2-hydroxy-N-[3-[(2-mercaptoethyl)amino]-3-oxopropyl]-3,3-dimethyl-4-(phosphonooxy)-,(2R)-](http://img.cochemist.com/ccimg/2300/2226-71-3_b.png)
![[2,2'-Bipyridine]-5,5'-dicarboxylic acid, 5,5'-diethyl ester](/data/chemimg/1612100/1762-46-5.png)
![[2,2'-Bipyridine]-5,5'-dicarboxylic acid, 5,5'-diethyl ester](/data/chemimg/1612100/1762-46-5_b.png)

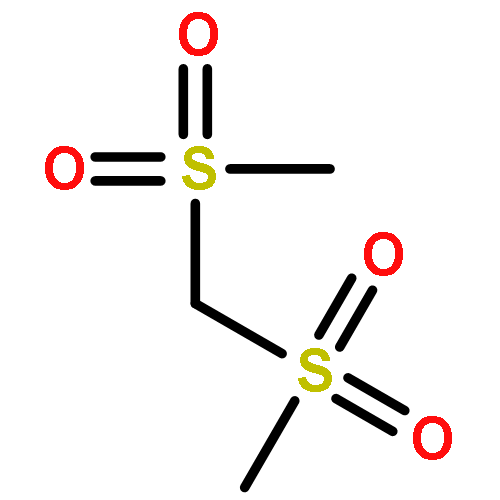


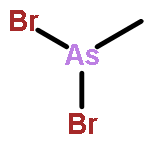
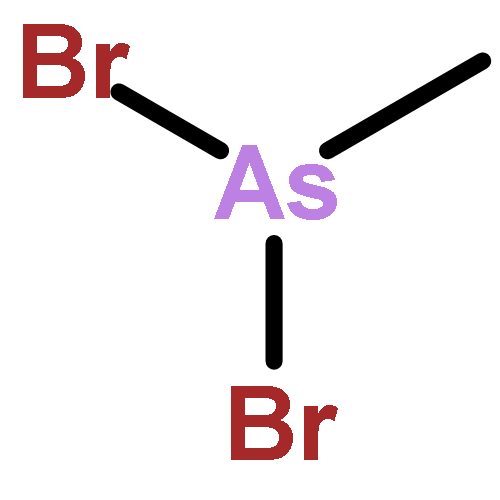
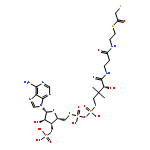
![(3AR,4R,5R,6AS)-4-FORMYL-2-OXOHEXAHYDRO-2H-CYCLOPENTA[B]FURAN-5-Y<WBR />L 4-BIPHENYLCARBOXYLATE](http://img.cochemist.com/ccimg/100/72-89-9.png)
![(3AR,4R,5R,6AS)-4-FORMYL-2-OXOHEXAHYDRO-2H-CYCLOPENTA[B]FURAN-5-Y<WBR />L 4-BIPHENYLCARBOXYLATE](http://img.cochemist.com/ccimg/100/72-89-9_b.png)