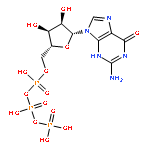
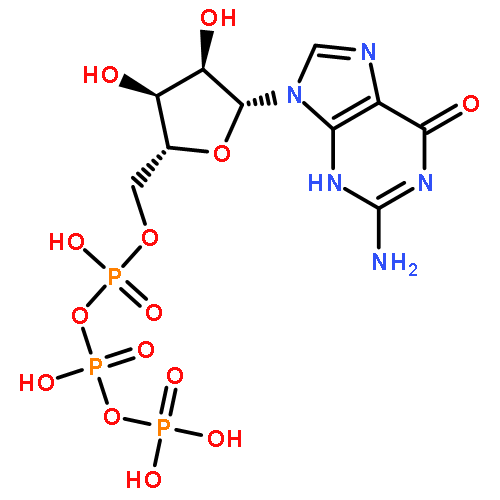
Co-reporter:Peter B. Moore
New Biotechnology 2017 Volume 38, Part A(Volume 38, Part A) pp:
Publication Date(Web):25 September 2017
DOI:10.1016/j.nbt.2016.07.010
In March 2014, a symposium was held at Aarhus University to commemorate the 40th anniversary of the founding of its Division of Biostructural Chemistry by Professor Brian Clark. This article is a lightly edited version of the talk the author gave to open that event. It consists of some reflections about the discipline known as structural biology, and some comments about the author’s interactions with Professor Clark and the Division of Biostructural Chemistry over the years.
Co-reporter:Peter B. Moore
PNAS 2013 Volume 110 (Issue 24 ) pp:9716-9721
Publication Date(Web):2013-06-11
DOI:10.1073/pnas.1307288110
Co-reporter:Peter B. Moore
Crystal Growth & Design 2011 Volume 11(Issue 3) pp:627
Publication Date(Web):January 19, 2011
DOI:10.1021/cg101282z
From ∼1970 to 2000, the year its crystal structure was solved, no biological structure was investigated more intensely than the ribosome. Some of the structural experiments done in the precrystallographic era produced information that significantly advanced our understanding of that enzyme. Other approaches yielded little that would be regarded as useful today, even if crystal structures had never been obtained. That experience is reviewed here to provide guidance for those faced with similar challenges in the future.
Co-reporter:PeterB. Moore
Israel Journal of Chemistry 2010 Volume 50( Issue 1) pp:36-44
Publication Date(Web):
DOI:10.1002/ijch.201000001
Abstract
The crystal structures that have been obtained for 23 different inhibitors bound to the large ribosomal subunit from Haloarcula marismortui are reviewed here. These structures provide important insights into how anti-ribosomal antibiotics inhibit protein synthesis, how species specificity arises, and the relationship between ribosomal mutations and antibiotic resistance. These structural studies also provide compelling evidence that the conformation of the peptidyl transferase center of the large ribosomal subunit is intrinsically variable, and that conformational equilibria play a role in determining its functional properties.
Co-reporter:Güliz Gürel, Gregor Blaha, Peter B. Moore, Thomas A. Steitz
Journal of Molecular Biology (29 May 2009) Volume 389(Issue 1) pp:146-156
Publication Date(Web):29 May 2009
DOI:10.1016/j.jmb.2009.04.005
Structures have been obtained for the complexes that tiamulin, homoharringtonine, and bruceantin form with the large ribosomal subunit of Haloarcula marismortui at resolutions ranging from 2.65 to 3.2 Å. They show that all these inhibitors block protein synthesis by competing with the amino acid side chains of incoming aminoacyl-tRNAs for binding in the A-site cleft in the peptidyl-transferase center, which is universally conserved. In addition, these structures support the hypothesis that the species specificity exhibited by the A-site cleft inhibitors is determined by the interactions they make, or fail to make, with a single nucleotide, U2504 (Escherichia coli). In the ribosome, the position of U2504 is controlled by its interactions with neighboring nucleotides, whose identities vary among kingdoms.
Co-reporter:Hong Jin, J. Patrick Loria, Peter B. Moore
Molecular Cell (27 April 2007) Volume 26(Issue 2) pp:205-215
Publication Date(Web):27 April 2007
DOI:10.1016/j.molcel.2007.03.014
Base pairing between the RNA components of box H/ACA small nucleolar ribonucleoproteins (snoRNPs) and sequences in other eukaryotic RNAs target specific uridines for pseudouridylation. An RNA called HJ1 has been developed that interacts with the rRNA sequence targeted by the 5′ pseudouridylation pocket of human U65 snoRNA the same way as intact U65 snoRNA. Sequences on both strands of the analog of the U65 snoRNP pseudouridylation pocket in HJ1 pair with its substrate sequence, and the resulting complex, called HJ3, is strongly stabilized by Mg2+. The solution structure of HJ3 reveals an Ω-shaped RNA interaction motif that has not previously been described, which is likely to be common to all box H/ACA snoRNP-substrate complexes. The topology of the complex explains why the access of substrate sequences to snoRNPs is facile and how uridine selection may occur when these complexes form.
Co-reporter:Peter B. Moore
Structure (14 October 2009) Volume 17(Issue 10) pp:1307-1315
Publication Date(Web):14 October 2009
DOI:10.1016/j.str.2009.08.015
The quality of many macromolecular crystal structures published recently has been enhanced through the use of new methods for treating the effects of molecular motion and disorder on diffraction patterns, among them a technique called translation, libration, screw-axis (TLS) parameterization. TLS parameterization rationalizes those effects in terms of domain-scale, rigid-body motions and, interestingly, the models for molecular motion that emerge when macromolecular diffraction data are analyzed this way often make sense biochemically. Here it is pointed out that all such models should be treated with caution until it is shown that they are consistent with the diffuse scatter produced by the crystals that provided the diffraction data from which they derive.
Co-reporter:Peter B. Moore
Structure (13 November 2007) Volume 15(Issue 11) pp:1350-1352
Publication Date(Web):13 November 2007
DOI:10.1016/j.str.2007.10.006
Co-reporter:Susan J. Schroeder, Gregor Blaha, Julian Tirado-Rives, Thomas A. Steitz, Peter B. Moore
Journal of Molecular Biology (13 April 2007) Volume 367(Issue 5) pp:1471-1479
Publication Date(Web):13 April 2007
DOI:10.1016/j.jmb.2007.01.081
Crystal structures of the 50 S ribosomal subunit from Haloarcula marismortui complexed with two antibiotics have identified new sites at which antibiotics interact with the ribosome and inhibit protein synthesis. 13-Deoxytedanolide binds to the E site of the 50 S subunit at the same location as the CCA of tRNA, and thus appears to inhibit protein synthesis by competing with deacylated tRNAs for E site binding. Girodazole binds near the E site region, but is somewhat buried and may inhibit tRNA binding by interfering with conformational changes that occur at the E site. The specificity of 13-deoxytedanolide for eukaryotic ribosomes is explained by its extensive interactions with protein L44e, which is an E site component of archaeal and eukaryotic ribosomes, but not of eubacterial ribosomes. In addition, protein L28, which is unique to the eubacterial E site, overlaps the site occupied by 13-deoxytedanolide, precluding its binding to eubacterial ribosomes. Girodazole is specific for eukarytes and archaea because it makes interactions with L15 that are not possible in eubacteria.
.jpg)
![Virginiamycin S1,4-[4-(dimethylamino)-N-methyl-L-phenylalanine]-5-[(2S,5R)-5-[[[(3S)-1-azabicyclo[2.2.2]oct-3-yl]thio]methyl]-4-oxo-2-piperidinecarboxylicacid]-](http://img.cochemist.com/ccimg/120200/120138-50-3.png)
![Virginiamycin S1,4-[4-(dimethylamino)-N-methyl-L-phenylalanine]-5-[(2S,5R)-5-[[[(3S)-1-azabicyclo[2.2.2]oct-3-yl]thio]methyl]-4-oxo-2-piperidinecarboxylicacid]-](http://img.cochemist.com/ccimg/120200/120138-50-3_b.png)
![3H-21,18-Nitrilo-1H,22H-pyrrolo[2,1-c][1,8,4,19]dioxadiazacyclotetracosine-1,7,16,22(4H,17H)-tetrone,8,9,14,15,24,25-hexahydro-14-hydroxy-4,12-dimethyl-3-(1-methylethyl)-, (3R,4R,5E,10E,12E,14S)-](http://img.cochemist.com/ccimg/21500/21411-53-0.png)
![3H-21,18-Nitrilo-1H,22H-pyrrolo[2,1-c][1,8,4,19]dioxadiazacyclotetracosine-1,7,16,22(4H,17H)-tetrone,8,9,14,15,24,25-hexahydro-14-hydroxy-4,12-dimethyl-3-(1-methylethyl)-, (3R,4R,5E,10E,12E,14S)-](http://img.cochemist.com/ccimg/21500/21411-53-0_b.png)
![Guanosine, N-methyl-6-O-[2-(4-nitrophenyl)ethyl]-](http://img.cochemist.com/ccimg/219700/219648-31-4.png)
![Guanosine, N-methyl-6-O-[2-(4-nitrophenyl)ethyl]-](http://img.cochemist.com/ccimg/219700/219648-31-4_b.png)


![2',3',5'-tris-O-acetyl-2-fluoro-O6-[2-(4-nitrophenyl)ethyl]inosine](http://img.cochemist.com/ccimg/171300/171284-48-3.png)
![2',3',5'-tris-O-acetyl-2-fluoro-O6-[2-(4-nitrophenyl)ethyl]inosine](http://img.cochemist.com/ccimg/171300/171284-48-3_b.png)
![Guanosine, 6-O-[2-(4-nitrophenyl)ethyl]-, 2',3',5'-triacetate](/data/chemimg/1156500/171284-47-2.png)
![Guanosine, 6-O-[2-(4-nitrophenyl)ethyl]-, 2',3',5'-triacetate](/data/chemimg/1156500/171284-47-2_b.png)
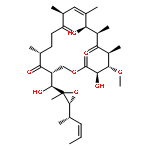
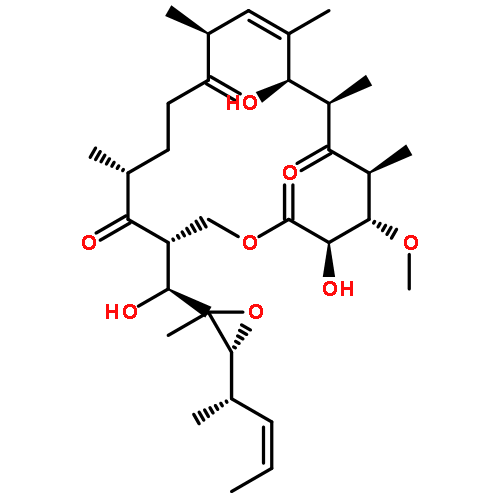


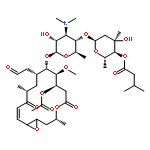
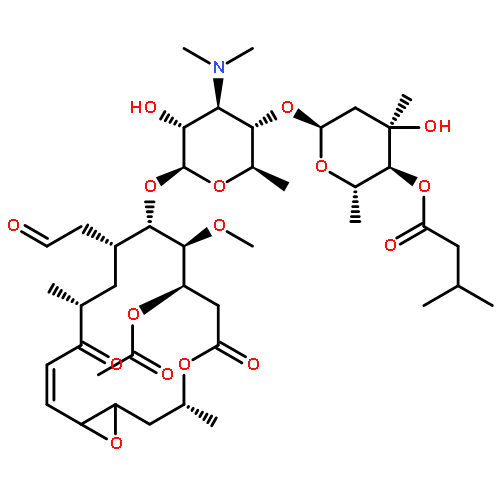

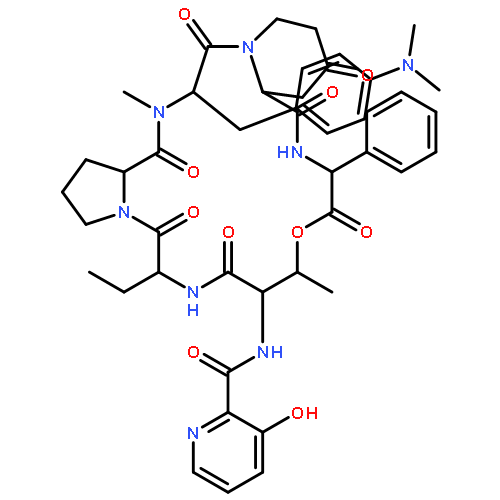
![b-D-erythro-Hex-2-enopyranuronicacid,4-[[3-amino-5-[(aminoiminomethyl)methylamino]-1-oxopentyl]amino]-1-(4-amino-2-oxo-1(2H)-pyrimidinyl)-1,2,3,4-tetradeoxy-,(S)-](http://img.cochemist.com/ccimg/2100/2079-00-7.png)
![b-D-erythro-Hex-2-enopyranuronicacid,4-[[3-amino-5-[(aminoiminomethyl)methylamino]-1-oxopentyl]amino]-1-(4-amino-2-oxo-1(2H)-pyrimidinyl)-1,2,3,4-tetradeoxy-,(S)-](http://img.cochemist.com/ccimg/2100/2079-00-7_b.png)


![2-Propenamide,N-[(1S)-1-(hydroxymethyl)-2-[(R)-[(methylthio)methyl]sulfinyl]ethyl]-3-(1,2,3,4-tetrahydro-6-methyl-2,4-dioxo-5-pyrimidinyl)-,(2E)-](http://img.cochemist.com/ccimg/1500/1404-64-4.png)
![2-Propenamide,N-[(1S)-1-(hydroxymethyl)-2-[(R)-[(methylthio)methyl]sulfinyl]ethyl]-3-(1,2,3,4-tetrahydro-6-methyl-2,4-dioxo-5-pyrimidinyl)-,(2E)-](http://img.cochemist.com/ccimg/1500/1404-64-4_b.png)


![1H-Imidazole-5-methanol,2-amino-a-[(1S)-2-amino-1-chloroethyl]-,hydrochloride (1:2), (aS)-](http://img.cochemist.com/ccimg/135900/135824-74-7.png)
![1H-Imidazole-5-methanol,2-amino-a-[(1S)-2-amino-1-chloroethyl]-,hydrochloride (1:2), (aS)-](http://img.cochemist.com/ccimg/135900/135824-74-7_b.png)
![D-glycero-L-gulo-Nonitol,4,8-anhydro-3,5-dideoxy-9-C-[[(2S)-2-hydroxy-2-[(2R,5R,6R)-tetrahydro-2-methoxy-5,6-dimethyl-4-methylene-2H-pyran-2-yl]acetyl]amino]-5,5-dimethyl-6-O-methyl-7,9-O-methylene-,(9S)-](http://img.cochemist.com/ccimg/115200/115185-92-7.png)
![D-glycero-L-gulo-Nonitol,4,8-anhydro-3,5-dideoxy-9-C-[[(2S)-2-hydroxy-2-[(2R,5R,6R)-tetrahydro-2-methoxy-5,6-dimethyl-4-methylene-2H-pyran-2-yl]acetyl]amino]-5,5-dimethyl-6-O-methyl-7,9-O-methylene-,(9S)-](http://img.cochemist.com/ccimg/115200/115185-92-7_b.png)


![2-[[(3,6-DIAMINO-5-HYDROXYHEXANOYL)AMINO]-METHYLAMINO]ACETIC ACID](http://img.cochemist.com/ccimg/33500/33404-78-3.png)
![2-[[(3,6-DIAMINO-5-HYDROXYHEXANOYL)AMINO]-METHYLAMINO]ACETIC ACID](http://img.cochemist.com/ccimg/33500/33404-78-3_b.png)






![3,4-Pyrrolidinediol,2-[(4-methoxyphenyl)methyl]-, 3-acetate, (2R,3S,4S)-](http://img.cochemist.com/ccimg/22900/22862-76-6.png)
![3,4-Pyrrolidinediol,2-[(4-methoxyphenyl)methyl]-, 3-acetate, (2R,3S,4S)-](http://img.cochemist.com/ccimg/22900/22862-76-6_b.png)