Co-reporter:Baoen Chen ; Fei Ye ; Lu Yu ; Guifang Jia ; Xiaotian Huang ; Xueju Zhang ; Shuying Peng ; Kai Chen ; Meining Wang ; Shouze Gong ; Ruihan Zhang ; Jinya Yin ; Haiyan Li ; Yiming Yang ; Hong Liu ; Jiwen Zhang ; Haiyan Zhang ; Ao Zhang ; Hualiang Jiang ; Cheng Luo
Journal of the American Chemical Society 2012 Volume 134(Issue 43) pp:17963-17971
Publication Date(Web):October 9, 2012
DOI:10.1021/ja3064149
The direct nucleic acid repair dioxygenase FTO is an enzyme that demethylates N6-methyladenosine (m6A) residues in mRNA in vitro and inside cells. FTO is the first RNA demethylase discovered that also serves a major regulatory function in mammals. Together with structure-based virtual screening and biochemical analyses, we report the first identification of several small-molecule inhibitors of human FTO demethylase. The most potent compound, the natural product rhein, which is neither a structural mimic of 2-oxoglutarate nor a chelator of metal ion, competitively binds to the FTO active site in vitro. Rhein also exhibits good inhibitory activity on m6A demethylation inside cells. These studies shed light on the development of powerful probes and new therapies for use in RNA biology and drug discovery.
Co-reporter:Jing Deng ; Ning Li ; Hongchuan Liu ; Zhili Zuo ; Oi Wah Liew ; Weijun Xu ; Gang Chen ; Xiankun Tong ; Wei Tang ; Jin Zhu ; Jianping Zuo ; Hualiang Jiang ; Cai-Guang Yang ; Jian Li ;Weiliang Zhu
Journal of Medicinal Chemistry 2012 Volume 55(Issue 14) pp:6278-6293
Publication Date(Web):June 28, 2012
DOI:10.1021/jm300146f
By virtual screening, compound 1 was found to be active against NS2B-NS3 protease (IC50 = 13.12 ± 1.03 μM). Fourteen derivatives (22) of compound 1 were synthesized, leading to the discovery of four new inhibitors with biological activity. In order to expand the chemical diversity of the inhibitors, small-molecule-based scaffold hopping was performed on the basis of the common scaffold of compounds 1 and 22. Twenty-one new compounds (23, 24) containing quinoline (new scaffold) were designed and synthesized. Protease inhibition assays revealed that 12 compounds with the new scaffold are inhibitors of NS2B-NS3 protease. Taken together, 17 new compounds were discovered as NS2B-NS3 protease inhibitors with IC50 values of 7.46 ± 1.15 to 48.59 ± 3.46 μM, and 8 compounds belonging to two different scaffolds are active to some extent against DENV based on luciferase reporter replicon-based assays. These novel chemical entities could serve as lead structures for discovering therapies against DENV.
Co-reporter:Chun-Xiao Song;Yao Sun;Dr. Qing Dai;Xing-Yu Lu;Miao Yu; Cai-Guang Yang; Chuan He
ChemBioChem 2011 Volume 12( Issue 11) pp:1682-1685
Publication Date(Web):
DOI:10.1002/cbic.201100278
Co-reporter:Baoen Chen, Hongchuan Liu, Xiaoxu Sun and Cai-Guang Yang
Molecular BioSystems 2010 vol. 6(Issue 11) pp:2143-2149
Publication Date(Web):11 Aug 2010
DOI:10.1039/C005148A
The human ABH2 and ABH3 proteins are functionally complementary in the oxidative demethylation of N1-methyl adenine (1-meA) and N3-methyl cytosine (3-meC) nucleotide bases. ABH3 displays higher activities with single-stranded DNA (ssDNA) in vitro, whereas ABH2 acts as the primary housekeeping enzyme in mammals for effectively repairing endogenously formed alkylated lesions in double-stranded DNA (dsDNA). Structurally, their overall protein folding is quite similar, but the most significant differences occur in the nucleotide recognition lid and the β-hairpin motif. We present here a site-directed mutational analysis and motif-swapping study to gain mechanistic insight into DNA substrate selection by ABH2 and ABH3. A V101A-F102A double mutant notably reduced ABH2 activity in dsDNA, indicating that this hydrophobic region appears to be important for damage searching and repair. The phenylalanine finger F102 is found to be crucial for ssDNA selection and repair as well; however, V101 shows reduced demethylating activity for only ssDNA and not dsDNA. The ABH2 R110A mutant completely loses the methyl base repair activity, suggesting that R110 is likely to be involved in the base flipping process. E175 and F124 contribute to nucleotide base specific selection and stabilization in the active site for repair. Additionally, swapping the RED residues in ABH3 to equivalent VFG residues in ABH2 endows ABH3 with activity in dsDNA repair as efficient as wild-type ABH2. Surprisingly, by changing just a few residues, the ABH3 protein can have very different selectivity towards ssDNA or dsDNA. This result indicates that the RED motif most likely prevents ABH3 binding and repair of dsDNA. Consistently, swapped ABH3 cross-links with dsDNA very well, confirming the determining roles of these residues in the initial DNA strand recognition. Overall, this work has provided a detailed understanding of the structural features of the ssDNA and dsDNA preferences of ABH2 and ABH3.

![2-Propen-1-ol, 3-[4-(1,1-dimethylethyl)phenyl]-, (E)-](http://img.cochemist.com/ccimg/117800/117721-77-4.png)
![2-Propen-1-ol, 3-[4-(1,1-dimethylethyl)phenyl]-, (E)-](http://img.cochemist.com/ccimg/117800/117721-77-4_b.png)
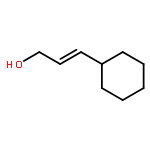
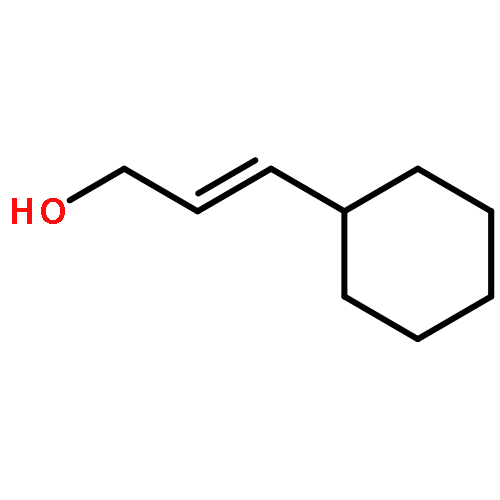
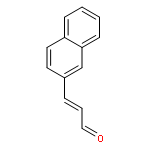
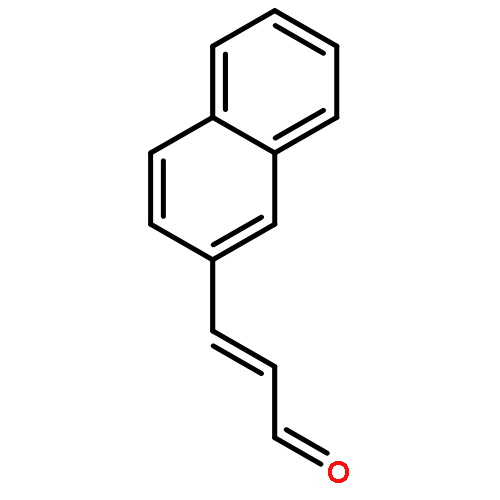
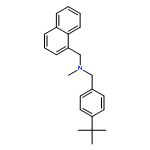
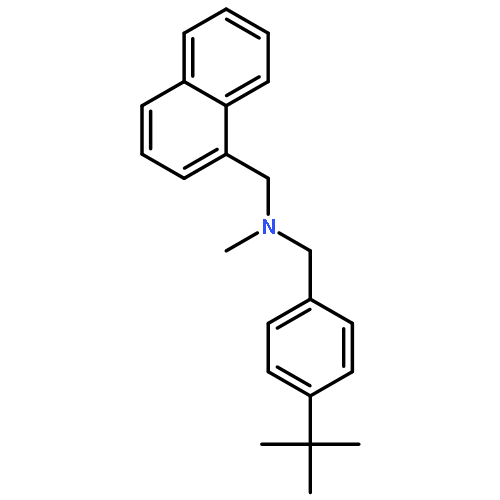
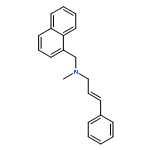
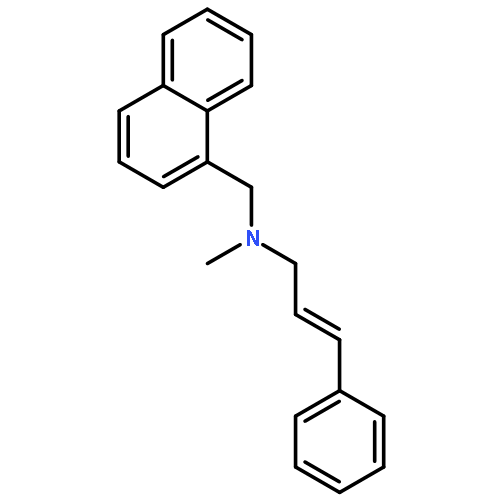
![1-Naphthalenemethanamine, N-[(2E)-3-phenyl-2-propenyl]-](http://img.cochemist.com/ccimg/92700/92610-10-1.png)
![1-Naphthalenemethanamine, N-[(2E)-3-phenyl-2-propenyl]-](http://img.cochemist.com/ccimg/92700/92610-10-1_b.png)
![2-Propenal, 3-[4-(1,1-dimethylethyl)phenyl]-, (2E)-](/data/chemimg/2010700/86604-07-1.png)
![2-Propenal, 3-[4-(1,1-dimethylethyl)phenyl]-, (2E)-](/data/chemimg/2010700/86604-07-1_b.png)
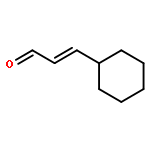
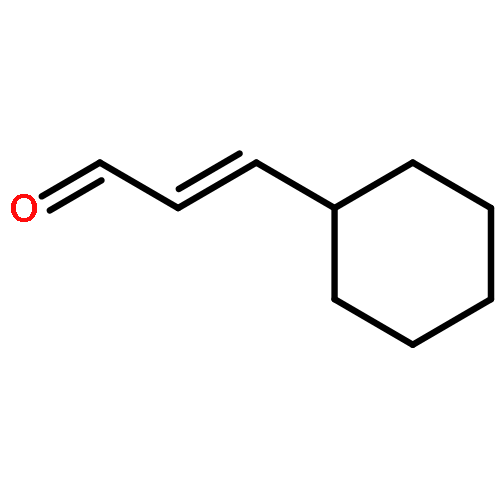
![3',6'-Dihydroxy-3H-spiro[isobenzofuran-1,9'-xanthen]-3-one](http://img.cochemist.com/ccimg/2400/2321-07-5.png)
![3',6'-Dihydroxy-3H-spiro[isobenzofuran-1,9'-xanthen]-3-one](http://img.cochemist.com/ccimg/2400/2321-07-5_b.png)