Co-reporter:Rurun Wang and Mohammad R. Seyedsayamdost
Organic Letters October 6, 2017 Volume 19(Issue 19) pp:
Publication Date(Web):September 18, 2017
DOI:10.1021/acs.orglett.7b02424
The secondary metabolome of the representative Roseobacter, Phaeobacter inhibens, was examined in response to algal sinapic acid. In addition to roseobacticides, sinapic acid induced the production of two new natural products, roseochelin A and B, which were characterized by NMR and X-ray crystallography. Functional assays showed that roseochelin B binds iron and is algaecidal against the algal host Emiliania huxleyi. It appears to be produced by a rarely observed combination of nonenzymatic and enzymatic transformations.
Co-reporter:Fei Xu, Behnam Nazari, Kyuho Moon, Leah B. Bushin, and Mohammad R. Seyedsayamdost
Journal of the American Chemical Society July 12, 2017 Volume 139(Issue 27) pp:9203-9203
Publication Date(Web):June 7, 2017
DOI:10.1021/jacs.7b02716
An important unresolved issue in microbial secondary metabolite production is the abundance of biosynthetic gene clusters that are not expressed under typical laboratory growth conditions. These so-called silent or cryptic gene clusters are sources of new natural products, but how they are silenced, and how they may be rationally activated are areas of ongoing investigation. We recently devised a chemogenetic high-throughput screening approach (“HiTES”) to discover small molecule elicitors of silent biosynthetic gene clusters. This method was successfully applied to a Gram-negative bacterium; it has yet to be implemented in the prolific antibiotic-producing streptomycetes. Herein we have developed a high-throughput transcriptional assay format in Streptomyces spp. by leveraging eGFP, inserted both at a neutral site and inside the biosynthetic cluster of interest, as a read-out for secondary metabolite synthesis. Using this approach, we successfully used HiTES to activate a silent gene cluster in Streptomyces albus J1074. Our results revealed the cytotoxins etoposide and ivermectin as potent inducers, allowing us to isolate and structurally characterize 14 novel small molecule products of the chosen cluster. One of these molecules is a novel antifungal, while several others inhibit a cysteine protease implicated in cancer. Studies addressing the mechanism of induction by the two elicitors led to the identification of a pathway-specific transcriptional repressor that silences the gene cluster under standard growth conditions. The successful application of HiTES will allow future interrogations of the biological regulation and chemical output of the countless silent gene clusters in Streptomyces spp.
Co-reporter:Behnam Nazari;Clarissa C. Forneris;Marcus I. Gibson;Kyuho Moon;Kelsey R. Schramma
MedChemComm (2010-Present) 2017 vol. 8(Issue 4) pp:780-788
Publication Date(Web):2017/04/20
DOI:10.1039/C6MD00637J
Glycopeptide antibiotics (GPAs) have served as potent clinical drugs and as an inspiration to chemists in various disciplines. Among known GPAs, complestatin, chloropeptin, and kistamicin are unique in that they contain an unusual indole–phenol crosslink. The mechanism of formation of this linkage is unknown, and to date, the biosynthetic gene cluster of only one GPA with an indole–phenol crosslink, that of complestatin, has been identified. Here, we report the genome sequence of the kistamicin producer Nonomuraea sp. ATCC 55076. We find that this strain harbours the largest actinobacterial chromosome to date, consisting of a single linear chromosome of ∼13.1 Mbp. AntiSMASH analysis shows that ∼32 biosynthetic gene clusters and ∼10% of the genome are devoted to production of secondary metabolites, which include 1,6-dihydroxyphenazine and nomuricin, a new anthraquinone-type pentacyclic compound that we report herein. The kistamicin gene cluster (kis) was identified bioinformatically. A unique feature of kis is that it contains two cytochrome P450 enzymes, which likely catalyze three crosslinking reactions. These findings set the stage for examining the biosynthesis of kistamicin and its unusual indole–phenol crosslink in the future.
Co-reporter:Dainan Mao;Leah B. Bushin;Kyuho Moon;Yihan Wu
PNAS 2017 114 (14 ) pp:E2920-E2928
Publication Date(Web):2017-04-04
DOI:10.1073/pnas.1619529114
Bacteria produce a diverse array of secondary metabolites that have been invaluable in the clinic and in research. These metabolites
are synthesized by dedicated biosynthetic gene clusters (BGCs), which assemble architecturally complex molecules from simple
building blocks. The majority of BGCs in a given bacterium are not expressed under normal laboratory growth conditions, and
our understanding of how they are silenced is in its infancy. Here, we have addressed this question in the Gram-negative model
bacterium Burkholderia thailandensis E264 using genetic, transcriptomic, metabolomic, and chemical approaches. We report that a previously unknown, quorum-sensing-controlled
LysR-type transcriptional regulator, which we name ScmR (for secondary metabolite regulator), serves as a global gatekeeper
of secondary metabolism and a repressor of numerous BGCs. Transcriptionally, we find that 13 of the 20 BGCs in B. thailandensis are significantly (threefold or more) up- or down-regulated in a scmR deletion mutant (ΔscmR). Metabolically, the ΔscmR strain displays a hyperactive phenotype relative to wild type and overproduces a number of compound families by 18- to 210-fold,
including the silent virulence factor malleilactone. Accordingly, the ΔscmR mutant is hypervirulent both in vitro and in a Caenorhabditis elegans model in vivo. Aside from secondary metabolism, ScmR also represses biofilm formation and transcriptionally activates ATP
synthesis and stress response. Collectively, our data suggest that ScmR is a pleiotropic regulator of secondary metabolism,
virulence, biofilm formation, and other stationary phase processes. A model for how the interplay of ScmR with pathway-specific
transcriptional regulators coordinately silences virulence factor production is proposed.
Co-reporter:Dainan Mao;Leah B. Bushin;Kyuho Moon;Yihan Wu
PNAS 2017 114 (14 ) pp:E2920-E2928
Publication Date(Web):2017-04-04
DOI:10.1073/pnas.1619529114
Bacteria produce a diverse array of secondary metabolites that have been invaluable in the clinic and in research. These metabolites
are synthesized by dedicated biosynthetic gene clusters (BGCs), which assemble architecturally complex molecules from simple
building blocks. The majority of BGCs in a given bacterium are not expressed under normal laboratory growth conditions, and
our understanding of how they are silenced is in its infancy. Here, we have addressed this question in the Gram-negative model
bacterium Burkholderia thailandensis E264 using genetic, transcriptomic, metabolomic, and chemical approaches. We report that a previously unknown, quorum-sensing-controlled
LysR-type transcriptional regulator, which we name ScmR (for secondary metabolite regulator), serves as a global gatekeeper
of secondary metabolism and a repressor of numerous BGCs. Transcriptionally, we find that 13 of the 20 BGCs in B. thailandensis are significantly (threefold or more) up- or down-regulated in a scmR deletion mutant (ΔscmR). Metabolically, the ΔscmR strain displays a hyperactive phenotype relative to wild type and overproduces a number of compound families by 18- to 210-fold,
including the silent virulence factor malleilactone. Accordingly, the ΔscmR mutant is hypervirulent both in vitro and in a Caenorhabditis elegans model in vivo. Aside from secondary metabolism, ScmR also represses biofilm formation and transcriptionally activates ATP
synthesis and stress response. Collectively, our data suggest that ScmR is a pleiotropic regulator of secondary metabolism,
virulence, biofilm formation, and other stationary phase processes. A model for how the interplay of ScmR with pathway-specific
transcriptional regulators coordinately silences virulence factor production is proposed.
Co-reporter:Yihan Wu, Mohammad R. Seyedsayamdost
Cell Chemical Biology 2017 Volume 24, Issue 12(Volume 24, Issue 12) pp:
Publication Date(Web):21 December 2017
DOI:10.1016/j.chembiol.2017.08.024
•HMNQ and HQNO, two natural quinolones, act synergistically to inhibit bacterial growth•Bacterial cytological profiles show they inhibit distinct steps in energy production•Primary metabolite analysis reveals that both also inhibit pyrimidine biosynthesis•Our results provide a functional rationale for the structural divergence in quinolonesMicrobial natural products are genetically encoded by dedicated biosynthetic gene clusters (BGCs). A given BGC usually produces a family of related compounds that share a core but contain variable substituents. Though common, the reasons underlying this divergent biosynthesis are in general unknown. Herein, we have addressed this issue using the hydroxyalkylquinoline (HAQ) family of natural products synthesized by Burkholderia thailandensis. Investigations into the detailed functions of two analogs show that they act synergistically in inhibiting bacterial growth. One analog is a nanomolar inhibitor of pyrimidine biosynthesis and at the same time disrupts the proton motive force. A second analog inhibits the cytochrome bc1 complex as well as pyrimidine biogenesis. These results provide a functional rationale for the divergent nature of HAQs. They imply that synergy and target promiscuity are driving forces for the evolution of tailoring enzymes that diversify the products of the HAQ biosynthetic pathway.Download high-res image (107KB)Download full-size image
Co-reporter:Bethany K. Okada, Yihan Wu, Dainan Mao, Leah B. Bushin, and Mohammad R. Seyedsayamdost
ACS Chemical Biology 2016 Volume 11(Issue 8) pp:2124
Publication Date(Web):June 23, 2016
DOI:10.1021/acschembio.6b00447
While bacterial genomes typically contain numerous secondary metabolite biosynthetic gene clusters, only a small fraction of these are expressed at any given time. The remaining majority is inactive or silent, and methods that awaken them would greatly expand our repertoire of bioactive molecules. We recently devised a new approach for identifying inducers of silent gene clusters and proposed that the clinical antibiotic trimethoprim acted as a global activator of secondary metabolism in Burkholderia thailandensis. Herein, we report that trimethoprim triggers the production of over 100 compounds that are not observed under standard growth conditions, thus drastically modulating the secondary metabolic output of B. thailandensis. Using MS/MS networking and NMR, we assign structures to ∼40 compounds, including a group of new molecules, which we call acybolins. With methods at hand for activation of silent gene clusters and rapid identification of small molecules, the hidden secondary metabolomes of bacteria can be interrogated.
Co-reporter:Rurun Wang;Maxwell Z. Wilson;Zemer Gitai
PNAS 2016 Volume 113 (Issue 6 ) pp:1630-1635
Publication Date(Web):2016-02-09
DOI:10.1073/pnas.1518034113
While we have come to appreciate the architectural complexity of microbially synthesized secondary metabolites, far less attention
has been paid to linking their structural features with possible modes of action. This is certainly the case with tropodithietic
acid (TDA), a broad-spectrum antibiotic generated by marine bacteria that engage in dynamic symbioses with microscopic algae.
TDA promotes algal health by killing unwanted marine pathogens; however, its mode of action (MoA) and significance for the
survival of an algal–bacterial miniecosystem remains unknown. Using cytological profiling, we herein determine the MoA of
TDA and surprisingly find that it acts by a mechanism similar to polyether antibiotics, which are structurally highly divergent.
We show that like polyether drugs, TDA collapses the proton motive force by a proton antiport mechanism, in which extracellular
protons are exchanged for cytoplasmic cations. The α-carboxy-tropone substructure is ideal for this purpose as the proton
can be carried on the carboxyl group, whereas the basicity of the tropylium ion facilitates cation export. Based on similarities
to polyether anticancer agents we have further examined TDA’s cytotoxicity and find it to exhibit potent, broad-spectrum anticancer
activities. These results highlight the power of MoA-profiling technologies in repurposing old drugs for new targets. In addition,
we identify an operon that confers TDA resistance to the producing marine bacteria. Bioinformatic and biochemical analyses
of these genes lead to a previously unknown metabolic link between TDA/acid resistance and the γ-glutamyl cycle. The implications
of this resistance mechanism in the context of the algal-bacterial symbiosis are discussed.
Co-reporter:Mohammad R. Seyedsayamdost ; Rurun Wang ; Roberto Kolter ;Jon Clardy
Journal of the American Chemical Society 2014 Volume 136(Issue 43) pp:15150-15153
Publication Date(Web):October 8, 2014
DOI:10.1021/ja508782y
Roseobacticides regulate the symbiotic relationship between a marine bacterium (Phaeobacter inhibens) and a marine microalga (Emiliania huxleyi). This relationship can be mutualistic, when the algal host provides food for the bacteria and the bacteria produce growth hormones and antibiotics for the algae, or parasitic, when the algae senesce and release p-coumaric acid. The released p-coumaric acid causes the bacteria to synthesize roseobacticides, which are nM−μM toxins for the algae. We examined the biosynthesis of roseobacticides and report that all roseobacticide precursors play critical roles during the mutualist phase of the symbiosis. Roseobacticides are biosynthesized from the algal growth promoter, the major food molecule provided by the algal cells, and the algal senescence signal that initiates the mutualist-to-parasite switch. Thus, molecules that are beneficial during mutualism are diverted to the synthesis of toxins during parasitism. A plausible mechanism for assembling roseobacticides from these molecules is proposed.
Co-reporter:Mohammad R. Seyedsayamdost
PNAS 2014 Volume 111 (Issue 20 ) pp:7266-7271
Publication Date(Web):2014-05-20
DOI:10.1073/pnas.1400019111
Over the past decade, bacterial genome sequences have revealed an immense reservoir of biosynthetic gene clusters, sets of
contiguous genes that have the potential to produce drugs or drug-like molecules. However, the majority of these gene clusters
appear to be inactive for unknown reasons prompting terms such as “cryptic” or “silent” to describe them. Because natural
products have been a major source of therapeutic molecules, methods that rationally activate these silent clusters would have
a profound impact on drug discovery. Herein, a new strategy is outlined for awakening silent gene clusters using small molecule
elicitors. In this method, a genetic reporter construct affords a facile read-out for activation of the silent cluster of
interest, while high-throughput screening of small molecule libraries provides potential inducers. This approach was applied
to two cryptic gene clusters in the pathogenic model Burkholderia thailandensis. The results not only demonstrate a prominent activation of these two clusters, but also reveal that the majority of elicitors
are themselves antibiotics, most in common clinical use. Antibiotics, which kill B. thailandensis at high concentrations, act as inducers of secondary metabolism at low concentrations. One of these antibiotics, trimethoprim,
served as a global activator of secondary metabolism by inducing at least five biosynthetic pathways. Further application
of this strategy promises to uncover the regulatory networks that activate silent gene clusters while at the same time providing
access to the vast array of cryptic molecules found in bacteria.
.jpg)
![1,2,3-Propanetricarboxylicacid, 2-hydroxy-,1-[2-[[(2S)-2-(2-carboxy-2-hydroxy-5-oxo-1-pyrrolidinyl)-1-oxopropyl]amino]ethyl]ester, (2R)-](http://img.cochemist.com/ccimg/157600/157568-17-7.png)
![1,2,3-Propanetricarboxylicacid, 2-hydroxy-,1-[2-[[(2S)-2-(2-carboxy-2-hydroxy-5-oxo-1-pyrrolidinyl)-1-oxopropyl]amino]ethyl]ester, (2R)-](http://img.cochemist.com/ccimg/157600/157568-17-7_b.png)
![N-[(3S,4R,4aR,6S)-3-(dichloromethyl)-1,6-dihydroxy-3-methyl-8-oxo-4,4a,5,6,7,8-hexahydro-3H-isochromen-4-yl]-L-alaninamide](http://img.cochemist.com/ccimg/74200/74141-69-8.png)
![N-[(3S,4R,4aR,6S)-3-(dichloromethyl)-1,6-dihydroxy-3-methyl-8-oxo-4,4a,5,6,7,8-hexahydro-3H-isochromen-4-yl]-L-alaninamide](http://img.cochemist.com/ccimg/74200/74141-69-8_b.png)
![(5S)-5-amino-N~2~-[(3S,4S,5S,6S)-3-(dichloromethyl)-1,5,6-trihydroxy-3-methyl-8-oxo-4,4a,5,6,7,8-hexahydro-3H-isochromen-4-yl]-4-oxo-L-norleucinamide](http://img.cochemist.com/ccimg/74200/74141-68-7.png)
![(5S)-5-amino-N~2~-[(3S,4S,5S,6S)-3-(dichloromethyl)-1,5,6-trihydroxy-3-methyl-8-oxo-4,4a,5,6,7,8-hexahydro-3H-isochromen-4-yl]-4-oxo-L-norleucinamide](http://img.cochemist.com/ccimg/74200/74141-68-7_b.png)
![PROPANAMIDE, 2-AMINO-N-[3-(DICHLOROMETHYL)-3,4,4A,5,6,7-HEXAHYDRO-5,6,8-TRIHYDROXY-3-METHYL-1-OXO-1H-2-BENZOPYRAN-4-YL]-, [3S-[3.ALPHA.,4.ALPHA.(R*),4A.BETA.,5.BETA.,6.ALPHA.]]-](http://img.cochemist.com/ccimg/72700/72615-20-4.png)
![PROPANAMIDE, 2-AMINO-N-[3-(DICHLOROMETHYL)-3,4,4A,5,6,7-HEXAHYDRO-5,6,8-TRIHYDROXY-3-METHYL-1-OXO-1H-2-BENZOPYRAN-4-YL]-, [3S-[3.ALPHA.,4.ALPHA.(R*),4A.BETA.,5.BETA.,6.ALPHA.]]-](http://img.cochemist.com/ccimg/72700/72615-20-4_b.png)












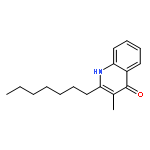
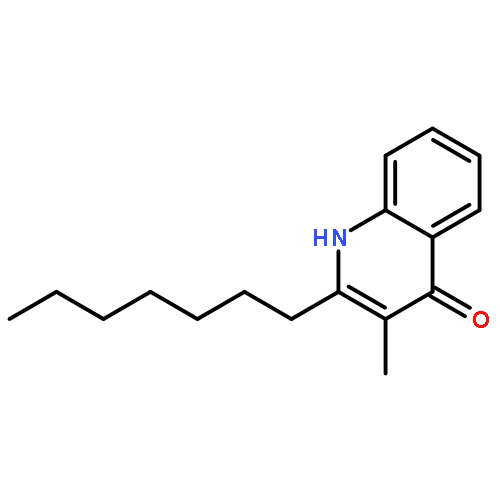
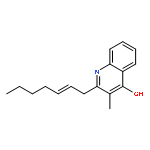
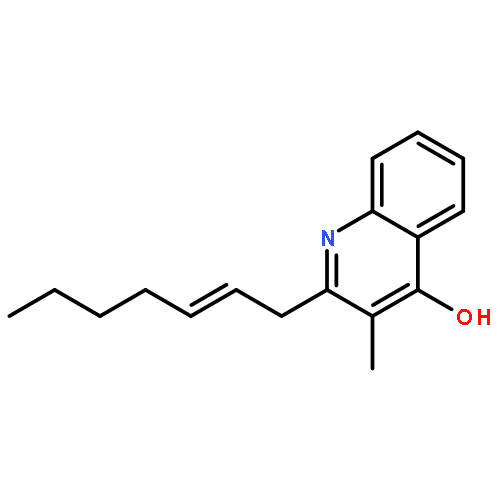


![[(3-chlorophenyl)hydrazono]malononitrile](http://img.cochemist.com/ccimg/600/555-60-2.png)
![[(3-chlorophenyl)hydrazono]malononitrile](http://img.cochemist.com/ccimg/600/555-60-2_b.png)