Co-reporter:Jiguo Qiu;Junjie Zhang;Yanting Zhang;Yuhong Wang;Lu Tong
Current Microbiology 2017 Volume 74( Issue 4) pp:508-514
Publication Date(Web):2017 April
DOI:10.1007/s00284-017-1205-2
We isolated a bacterial strain JQ135 from municipal wastewater, which was capable of efficiently degrading picolinic acid (PA). Based on the physico-biochemical characteristics and 16S rDNA analysis, strain JQ135 was identified as Alcaligenes faecalis. In addition, strain JQ135 produced an orange pigment when cultured in the Luria-Bertani medium, which is different from the previously reported strains of A. faecalis. During the degradation of PA by the resting strain JQ135 cells, only one intermediate, 6-hydroxypicolinic acid (6HPA), was detected by ultraviolet spectrophotometry, high-pressure liquid chromatography, and liquid chromatography–mass spectrometry. A random transposon mutagenesis library of strain JQ135 was constructed. One mutant, Mut-G31, could convert PA into 6HPA without further degradation. The disrupted gene (orf2) was amplified from Mut-G31, and its product showed 32% identity to the 3-deoxy-d-manno-octulosonic acid kinase (KdkA) from Haemophilus influenzae. Results from complementation analysis confirmed that GTG was the initiation codon of the kdkA-like orf2, and that it was essential for PA biodegradation by strain JQ135. This study provides the first genetic evidence for the bacterial degradation of PA.
Co-reporter:Cui-Wei Chu;Qing Chen;Cheng-Hong Wang;Hong-Mei Wang
Antonie van Leeuwenhoek 2016 Volume 109( Issue 5) pp:611-618
Publication Date(Web):2016 May
DOI:10.1007/s10482-016-0664-y
A Gram-negative, aerobic, short rod-shaped, pink-pigmented, non-motile bacterium, designated BUT-13T, was isolated from activated sludge of an herbicide-manufacturing wastewater treatment facility in Jiangsu province, China. Growth was observed at 0–5.5 % NaCl, pH 6.0–9.0 and 12–37 °C. Phylogenetic analysis based on 16S rRNA gene sequences indicated that strain BUT-13T is a member of the genus Roseomonas, and shows high sequence similarities to R. pecuniae N75T (98.0 %) and R. rosea 173-96T (97.5 %), and lower (<97 %) sequence similarities to all other Roseomonas species. Chemotaxonomic analysis revealed that strain BUT-13T possesses Q-10 as the predominant ubiquinone; summed feature 8 (C18:1w7c and/or C18:1w6c; 38.8 %), C18:0 (16.6 %), C16:0 (15.2 %), summed feature 3 (C16:1ω6c and/or C16:1ω7; 7.9 %) and C18:1w9c (4.7 %) as the major fatty acids. The polar lipids were found to consist of two aminolipids, a glycolipid, a phospholipid, a phosphoglycolipid, phosphatidylcholine, phosphatidylethanolamine and diphosphatidylglycerol. Strain BUT-13T showed low DNA–DNA relatedness with R. pecuniae N75T (45.2 %) and R. rosea 173-96T (51.2 %). The DNA G+C content was determined to be 67.6 mol%. Based on the phylogenetic analysis, DNA–DNA hybridization and chemotaxonomic analysis, as well as biochemical characteristics, strain BUT-13T can be clearly distinguished from all currently recognised Roseomonas species and should be classified as a novel species of the genus Roseomonas, for which the name Roseomonas chloroacetimidivorans sp. nov. is proposed. The type strain is BUT-13T (CCTCC AB 2015299T = JCM 31050T).
Co-reporter:Jiguo Qiu;Na Li;Zhenmei Lu;Youjian Yang
Applied Microbiology and Biotechnology 2016 Volume 100( Issue 23) pp:10019-10029
Publication Date(Web):2016 December
DOI:10.1007/s00253-016-7805-0
Nornicotine is a natural alkaloid produced by plants in the genus Nicotiana and is structurally related to nicotine. Importantly, nornicotine is the direct precursor of tobacco-specific nitrosamine N′-nitrosonornicotine, which is a highly potent human carcinogen. Microbial detoxification and degradation of nicotine have been well characterized; however, until now, there has been no information on the molecular mechanism of nornicotine degradation. In this study, we demonstrate the transformation of nornicotine by the nicotine-degrading strain Shinella sp. HZN7. Three transformation products were identified as 6-hydroxy-nornicotine, 6-hydroxy-myosmine, and 6-hydroxy-pseudooxy-nornicotine by UV spectroscopy, high-resolution mass spectrometry, nuclear magnetic resonance, and Fourier transform-infrared spectroscopy analyses. The two-component nicotine dehydrogenase genes nctA1 and nctA2 were cloned, and their product, NctA, was confirmed to be responsible for the conversion of nornicotine into 6-hydroxy-nornicotine as well as nicotine into 6-hydroxy-nicotine. The 6-hydroxy-nicotine oxidase, NctB, catalyzed the oxidation of 6-hydroxy-nornicotine to 6-hydroxy-myosmine, and it spontaneously hydrolyzed into 6-hydroxy-pseudooxy-nornicotine. However, 6-hydroxy-pseudooxy-nornicotine could not be further degraded by strain HZN7. This study demonstrated that nornicotine is partially transformed by strain HZN7 via nicotine degradation pathway.
Co-reporter:Zhi-Juan Nie, Bao-Jian Hang, Shu Cai, Xiang-Ting Xie, Jian He, and Shun-Peng Li
Journal of Agricultural and Food Chemistry 2011 Volume 59(Issue 11) pp:6040-6046
Publication Date(Web):May 2, 2011
DOI:10.1021/jf200397t
Cyhalofop-butyl (CyB) is a widely used aryloxyphenoxy propanoate (AOPP) herbicide for control of grasses in rice fields. Five CyB-degrading strains were isolated from rice field soil and identified as Agromyces sp., Stenotrophomonas sp., Aquamicrobium sp., Microbacterium sp., and Pseudomonas azotoformans; the results revealed high biodiversity of CyB-degrading bacteria in rice soil. One strain, P. azotoformans QDZ-1, degraded 84.5% of 100 mg L–1 CyB in 5 days of incubation in a flask and utilized CyB as carbon source for growth. Strain QDZ-1 could also degrade a wide range of other AOPP herbicides. An esterase gene, chbH, which hydrolyzes CyB to cyhalofop acid (CyA), was cloned from strain QDZ-1 and functionally expressed. A chbH-disrupted mutant dchbH was constructed by insertion mutation. Mutant dchbH could not degrade and utilize CyB, suggesting that chbH was the only esterase gene responsible for CyB degradation in strain QDZ-1. ChbH hydrolyzed all AOPP herbicides tested as well as permethrin. The catalytic efficiency of ChbH toward different AOPP herbicides followed the order quizalofop-P-ethyl ≈ fenoxaprop-P-ethyl > CyB ≈ fluazifop-P-butyl > diclofop-methyl ≈ haloxyfop-P-methyl; the results indicated that the chain length of the alcohol moiety strongly affected the biodegradability of the AOPP herbicides, whereas the substitutions in the aromatic ring had only a slight influence.
Co-reporter:Jun Zhang, Jin-Wei Zheng, Bin Liang, Cheng-Hong Wang, Shu Cai, Ying-Ying Ni, Jian He, and Shun-Peng Li
Journal of Agricultural and Food Chemistry 2011 Volume 59(Issue 9) pp:4614-4621
Publication Date(Web):March 21, 2011
DOI:10.1021/jf104695g
A butachlor-degrading strain, designated FLY-8, was isolated from rice field soil and was identified as Paracoccus sp. Strain FLY-8 could degrade and utilize six chloroacetamide herbicides as carbon sources for growth, and the degradation rates followed the order alachlor > acetochlor > propisochlor > butachlor > pretilachlor > metolachlor. The influence of molecular structure of the chloroacetamide herbicides on the microbial degradation rate was first analyzed; the results indicated that the substitutions of alkoxymethyl side chain with alkoxyethyl side chain greatly reduced the degradation efficiencies; the length of amide nitrogen’s alkoxymethyl significantly affected the biodegradability of these herbicides: the longer the alkyl was, the slower the degradation efficiencies occurred. The phenyl alkyl substituents have no obvious influence on the degradation efficiency. The pathway of butachlor complete mineralization was elucidated on the basis of the results of metabolite identification and enzyme assays. Butachlor was degraded to alachlor by partial C-dealkylation and then converted to 2-chloro-N-(2,6-dimethylphenyl)acetamide by N-dealkylation, which subsequently transformed to 2,6-diethylaniline, which was further degraded via the metabolites aniline and catechol, and catechol was oxidized through an ortho-cleavage pathway. This study highlights an important potential use of strain FLY-8 for the in situ bioremediation of chloroacetamide herbicides and their metabolite-contaminated environment.
Co-reporter:Bao-zhan Wang;Yun Ma;Wei-you Zhou
World Journal of Microbiology and Biotechnology 2011 Volume 27( Issue 10) pp:2315-2324
Publication Date(Web):2011 October
DOI:10.1007/s11274-011-0698-2
A synthetic pyrethroid (SP)-degrading bacterium, designated pyd-1, was isolated from SPcontaminated soil. Based on its phenotypic and genotypic properties, the strain was identified as Ochrobactrum tritici. Strain pyd-1 was able to degrade a wide range of SPs, and its degradation efficiencies were dependent on the molecular structure of the SP. Interestingly, the strain degraded cis- and trans-permethrin (cypermethrin) at nearly the same rate and possessed approximately equal hydrolysis activities toward the two enantiomers of fenpropathrin. These results suggest that different isomers of SPs are degraded with equal efficiency by strain pyd-1. We studied the metabolic pathway of fenpropathrin degradation in strain pyd-1 by metabolite identification and enzymatic analysis. Fenpropathrin is degraded by hydrolysis of the carboxylester linkage to yield 2,2,3,3-tetramethylcyclopropanecarboxylic acid and 3-phenoxybenzaldehyde, which is converted to 3-phenoxybenzoic acid (PBA). PBA is further metabolized to 4-hydroxy-3-phenoxybenzoic acid (4-hydroxy-PBA). 4-Hydroxy-PBA is oxidized to protocatechuate and p-hydroquinone. Protocatechuate is further oxidized through an ortho-cleavage pathway, and p-hydroquinone is degraded via 1,2,4-benzenetriol.
Co-reporter:Zhi-Fei Lang;Jing-Jing Shen;Shu Cai;Jun Zhang
Current Microbiology 2011 Volume 63( Issue 2) pp:145-150
Publication Date(Web):2011 August
DOI:10.1007/s00284-011-9953-x
A multiple herbicide-resistant acetohydroxyacid synthase (rAHAS) gene was cloned from Pseudomonas sp. Lm10. Sequence analysis showed that the rAHAS regulatory subunit was identical to that of Pseudomonasputida KT2440 (sensitive AHAS, sAHAS), whereas six different sites [H134→N (rAHAS→sAHAS), A135→P, S136→T, I210→V, F264→Y, and S486→W] were found in the catalytic subunit. The rAHAS and sAHAS were over expressed, purified and characterized. rAHAS showed higher resistance to four kinds of AHAS-inhibitor herbicides than sAHAS. The resistance factor of rAHAS was 56.0-fold, 12.6-fold, 6.5-fold, and 9.2-fold as compared with sAHAS when metsulfuron-methyl, imazethapyr, flumetsulam, and pyriminobac-methyl used as inhibitor, respectively. The specific activity of rAHAS was lower than that of sAHAS and the Km value of rAHAS for pyruvate was approximately onefold higher than the corresponding value for sAHAS. Data from site-directed mutagenesis demonstrated that alteration at A135, F264, and S486 resulted in resistance reduction, while the mutation at H134, S136, and I210 has little effect on the resistance. A135 was mainly responsible for resistance to imidazolinone; F264 conferred resistance to sulfonylurea and triazolopyrimidine sulfonamide; and S486 showed multiple herbicides resistance to the four herbicides.
Co-reporter:Haiyan Ni, Li Yao, Na Li, Qin Cao, ... Jian He
Journal of Environmental Sciences (March 2016) Volume 41() pp:121-127
Publication Date(Web):1 March 2016
DOI:10.1016/j.jes.2015.04.035
A bacterium strain Y3, capable of efficiently degrading pendimethalin, was isolated from activated sludge and identified as Bacillus subtilis according to its phenotypic features and 16S rRNA phylogenetic analysis. This strain could grow on pendimethalin as a sole carbon source and degrade 99.5% of 100 mg/L pendimethalin within 2.5 days in batch liquid culture, demonstrating a greater efficiency than any other reported strains. Three metabolic products, 6-aminopendimethalin, 5-amino-2-methyl-3-nitroso-4-(pentan-3-ylamino) benzoic acid, and 8-amino-2-ethyl-5-(hydroxymethyl)-1,2-dihydroquinoxaline-6-carboxylic acid, were identified by HPLC-MS/MS, and a new microbial degradation pathway was proposed. A nitroreductase catalyzing nitroreduction of pendimethalin to 6-aminopendimethalin was detected in the cell lysate of strain Y3. The cofactor was nicotinamide adenine dinucleotide phosphate (NADPH) or more preferably nicotinamide adenine dinucleotide (NADH). The optimal temperature and pH for the nitroreductase were 30°C and 7.5, respectively. Hg2 +, Ni2 +, Pb2 +, Co2 +, Mn2 + Cu2 +, Ag+, and EDTA severely inhibited the nitroreductase activity, whereas Fe2 +, Mg2 +, and Ca2 + enhanced it. This study provides an efficient pendimethalin-degrading microorganism and broadens the knowledge of the microbial degradation pathway of pendimethalin.Download full-size image
Co-reporter:Peng Guo, Baozhan Wang, BaoJian Hang, Lian Li, Shinawar Waseem Ali, Jian He, Shunpeng Li
International Biodeterioration & Biodegradation (December 2009) Volume 63(Issue 8) pp:1107-1112
Publication Date(Web):1 December 2009
DOI:10.1016/j.ibiod.2009.09.008
A pyrethroid-degrading bacterium strain JZ-2 was isolated from activated sludge treating pyrethroid-manufacturing wastewater. Based on the morphological, physiological and biochemical characterization, and phylogenetic analysis of the 16S rRNA gene sequence, the strain was identified as Sphingobium sp. Strain JZ-2 was capable of degrading fenpropathrin, cypermethrin, permethrin, cyhalothrin, deltamethrin, fenvalerate and bifenthrin. This strain degraded fenpropathrin by hydrolysis of the carboxylester linkage to yield 3-phenoxybenzaldehyde and 2,2,3,3-tetramethylcyclopropanecarboxylic acid. 3-Phenoxybenzaldehyde, 3-phenoxybenzoate, protocatechuate and catechol are the intermediates of fenpropathrin degradation. Protocatechuate and catechol were further oxidized by ortho-cleavage pathway. A novel pyrethroid hydrolase from cell-free extract was purified 108.5-fold to apparent homogeneity with a 10.2% overall recovery. It was a monomer with a molecular mass of 31 ± 1 kDa, a pI of 4.85. The optimal pH and temperature were 7.5 and 40 °C, respectively. No cofactors or coenzymes were required for the pyrethroid-hydrolysis activity. The enzyme was strongly inhibited by many irons (Ag+, Cu2+, Hg2+ and Zn2+), SDS, p-chloromercuribenzoic acid, phenylmethylsulfonyl fluoride and malathion.

![cyano(3-phenoxyphenyl)methyl (1S,3S)-3-[(1Z)-2-chloro-3,3,3-trifluoroprop-1-en-1-yl]-2,2-dimethylcyclopropanecarboxylate](http://img.cochemist.com/ccimg/68100/68085-85-8.png)
![cyano(3-phenoxyphenyl)methyl (1S,3S)-3-[(1Z)-2-chloro-3,3,3-trifluoroprop-1-en-1-yl]-2,2-dimethylcyclopropanecarboxylate](http://img.cochemist.com/ccimg/68100/68085-85-8_b.png)


![[3-(phosphonomethyl)phenyl]methylphosphonic Acid](http://img.cochemist.com/ccimg/144100/144052-40-4.png)
![[3-(phosphonomethyl)phenyl]methylphosphonic Acid](http://img.cochemist.com/ccimg/144100/144052-40-4_b.png)


![2-({[(2,4-DIMETHYLPHENOXY)ACETYL]CARBAMOTHIOYL}AMINO)BENZOIC ACID](http://img.cochemist.com/ccimg/78200/78194-29-3.png)
![2-({[(2,4-DIMETHYLPHENOXY)ACETYL]CARBAMOTHIOYL}AMINO)BENZOIC ACID](http://img.cochemist.com/ccimg/78200/78194-29-3_b.png)
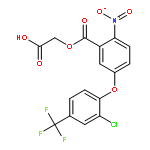
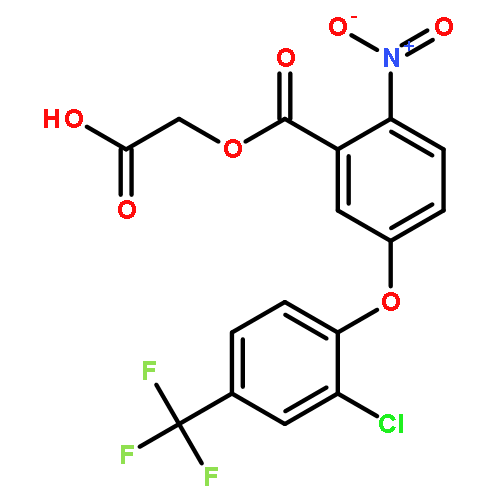






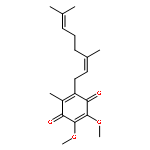
![TETRAETHYL [1,3-PHENYLENEBIS(METHYLENE)]BIS(PHOSPHONATE)](http://img.cochemist.com/ccimg/56900/56875-38-8.png)
![TETRAETHYL [1,3-PHENYLENEBIS(METHYLENE)]BIS(PHOSPHONATE)](http://img.cochemist.com/ccimg/56900/56875-38-8_b.png)
![(1R,7AR)-7-{[(2,3-DIHYDROXY-2-ISOPROPYLBUTANOYL)OXY]METHYL}-2,3,5,7A-TETRAHYDRO-1H-PYRROLIZIN-1-YL (2E)-2-METHYL-2-BUTENOATE](http://img.cochemist.com/ccimg/42100/42092-05-7.png)
![(1R,7AR)-7-{[(2,3-DIHYDROXY-2-ISOPROPYLBUTANOYL)OXY]METHYL}-2,3,5,7A-TETRAHYDRO-1H-PYRROLIZIN-1-YL (2E)-2-METHYL-2-BUTENOATE](http://img.cochemist.com/ccimg/42100/42092-05-7_b.png)

















![[4-(phosphonomethyl)phenyl]methylphosphonic Acid](http://img.cochemist.com/ccimg/4600/4546-06-9.png)
![[4-(phosphonomethyl)phenyl]methylphosphonic Acid](http://img.cochemist.com/ccimg/4600/4546-06-9_b.png)








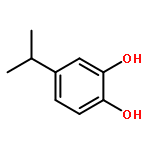
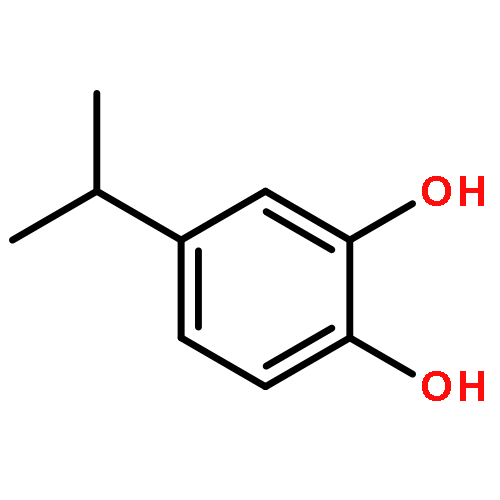






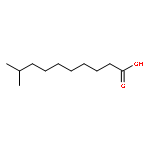









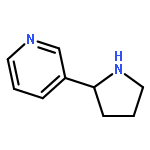

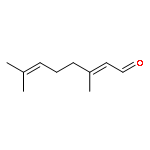
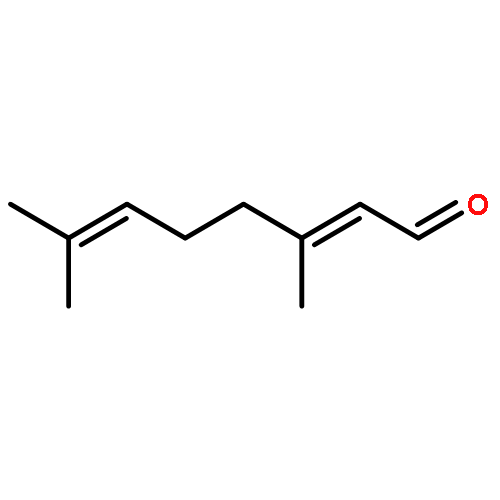


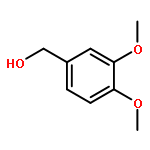
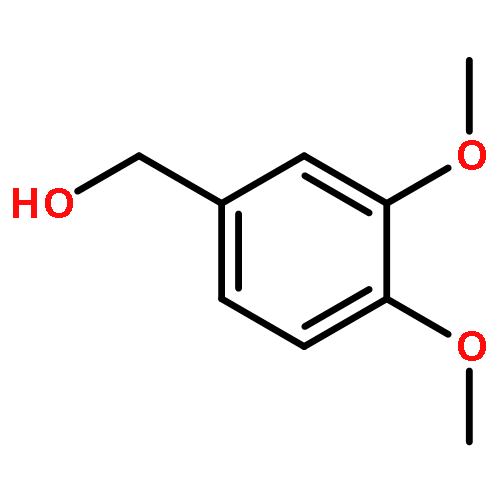




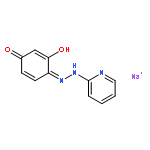
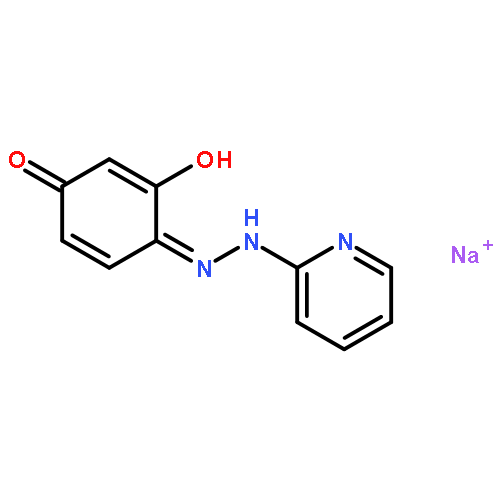
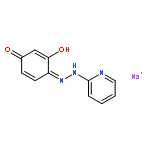
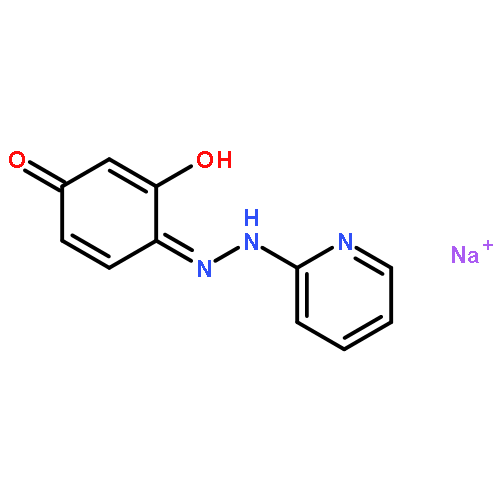
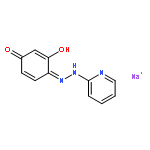
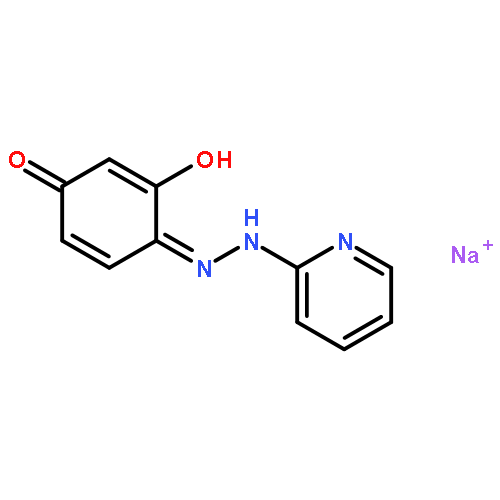




















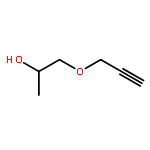
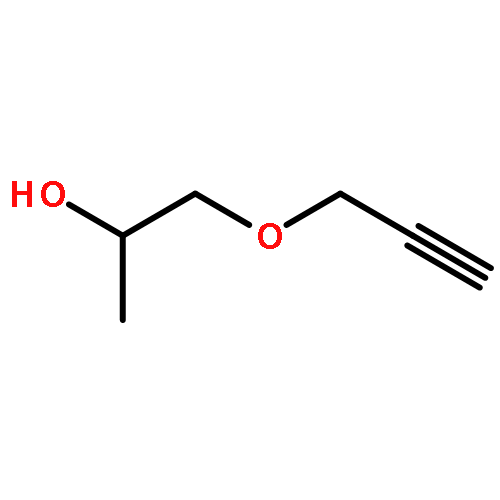




![4-[(1E)-1-{4-[2-(DIMETHYLAMINO)ETHOXY]PHENYL}-2-PHENYL-1-BUTEN-1-<WBR />YL]PHENYL ACETATE](http://img.cochemist.com/ccimg/2400/2394-68-5.png)
![4-[(1E)-1-{4-[2-(DIMETHYLAMINO)ETHOXY]PHENYL}-2-PHENYL-1-BUTEN-1-<WBR />YL]PHENYL ACETATE](http://img.cochemist.com/ccimg/2400/2394-68-5_b.png)