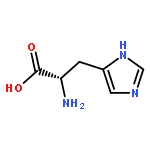
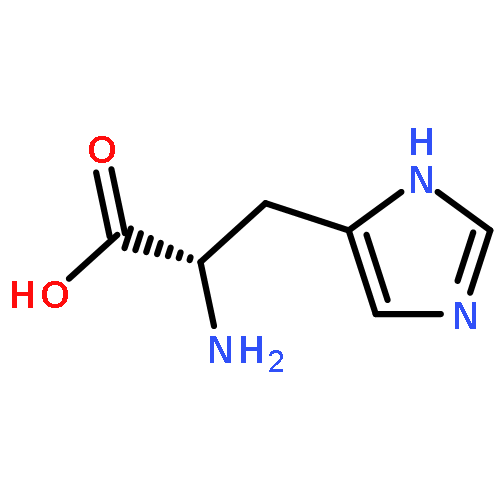
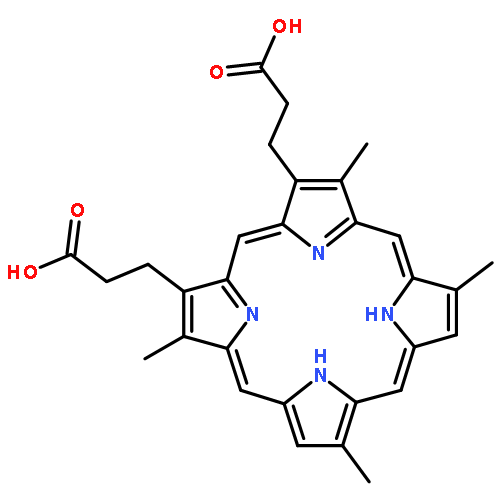
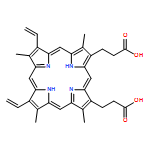
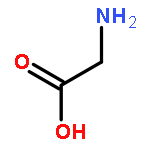
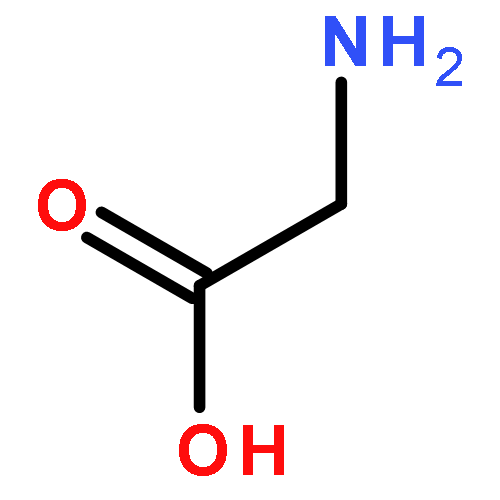
Co-reporter:Joseph W. LaMattina;David B. Nix;William Nicholas Lanzilotta
PNAS 2016 Volume 113 (Issue 43 ) pp:12138-12143
Publication Date(Web):2016-10-25
DOI:10.1073/pnas.1603209113
All of the heme-degrading enzymes that have been characterized to date require molecular oxygen as a cosubstrate. Escherichia coli O157:H7 has been shown to express heme uptake and transport proteins, as well as use heme as an iron source. This enteric
pathogen colonizes the anaerobic space of the lower intestine in mammals, yet no mechanism for anaerobic heme degradation
has been reported. Herein we provide evidence for an oxygen-independent heme-degradation pathway. Specifically, we demonstrate
that ChuW is a radical S-adenosylmethionine methyltransferase that catalyzes a radical-mediated mechanism facilitating iron liberation and the production
of the tetrapyrrole product we termed “anaerobilin.” We further demonstrate that anaerobilin can be used as a substrate by
ChuY, an enzyme that is coexpressed with ChuW in vivo along with the heme uptake machinery. Our findings are discussed in
terms of the competitive advantage this system provides for enteric bacteria, particularly those that inhabit an anaerobic
niche in the intestines.
Co-reporter:Jonathan M. Demick and William N. Lanzilotta
Biochemistry 2011 Volume 50(Issue 4) pp:
Publication Date(Web):December 23, 2010
DOI:10.1021/bi101255e
Activation of glycyl radical enzymes (GREs) by S-adenosylmethonine (AdoMet or SAM)-dependent enzymes has long been shown to proceed via the reductive cleavage of SAM. The AdoMet-dependent (or radical SAM) enzymes catalyze this reaction by using a [4Fe-4S] cluster to reductively cleave AdoMet to form a transient 5′-deoxyadenosyl radical and methionine. This radical is then transferred to the GRE, and methionine and 5′-deoxyadenosine are also formed. In contrast to this paradigm, we demonstrate that generation of a glycyl radical on the B12-independent glycerol dehydratase by the glycerol dehydratase activating enzyme results in formation of 5′-deoxy-5′-(methylthio)adenosine and not 5′-deoxyadenosine. This demonstrates for the first time that radical SAM activases are also capable of an alternative cleavage pathway for SAM.
Co-reporter:Bret D. Dillard;Jonathan M. Demick
JBIC Journal of Biological Inorganic Chemistry 2011 Volume 16( Issue 6) pp:949-959
Publication Date(Web):2011 August
DOI:10.1007/s00775-011-0795-6
High-resolution crystal structures of Pyrococcus furiosus rubrerythrin (PfRbr) in the resting (all-ferrous) state and at time points following exposure of the crystals to hydrogen peroxide are reported. This approach was possible because of the relativity slow turnover of PfRbr at room temperature. To this end, we were able to perform time-dependent peroxide treatment of the fully reduced enzyme, under strictly anaerobic conditions, in the crystalline state. In this work we demonstrate, for the first time, that turnover of a thermophilic rubrerythrin results in approximately 2-Å movement of one iron atom in the diiron site from a histidine to a carboxylate ligand. These results confirm that, despite the domain-swapped architecture, the hyperthermophilic rubrerythrins also utilize the classic combination of iron sites together with redox-dependent iron toggling to selectively reduce hydrogen peroxide over dioxygen. In addition, we have identified previously unobserved intermediates in the reaction cycle and observed structural changes that may explain the enzyme precipitation observed for the all-iron form of PfRbr upon oxidation to the all-ferric state.
Co-reporter:Amy Medlock;Larkin Swartz;Tamara A. Dailey;Harry A. Dailey;William N. Lanzilotta;
Proceedings of the National Academy of Sciences 2007 104(6) pp:1789-1793
Publication Date(Web):January 29, 2007
DOI:10.1073/pnas.0606144104
Ferrochelatase, the terminal enzyme in heme biosynthesis, catalyzes the insertion of ferrous iron into protoporphyrin IX to
form protoheme IX. Human ferrochelatase is a homodimeric, inner mitochondrial membrane-associated enzyme that possesses an
essential [2Fe-2S] cluster. In this work, we report the crystal structure of human ferrochelatase with the substrate protoporphyrin
IX bound as well as a higher resolution structure of the R115L variant without bound substrate. The data presented reveal
that the porphyrin substrate is bound deep within an enclosed pocket. When compared with the location of N-methylmesoporphyrin in the Bacillus subtilis ferrochelatase, the porphyrin is rotated by ≈100° and is buried an additional 4.5 Å deeper within the active site. The propionate
groups of the substrate do not protrude into solvent and are bound in a manner similar to what has been observed in uroporphyrinogen
decarboxylase. Furthermore, in the substrate-bound form, the jaws of the active site mouth are closed so that the porphyrin
substrate is completely engulfed in the pocket. These data provide insights that will aid in the determination of the mechanism
for ferrochelatase.
Co-reporter:Amy E. Medlock, Michael Carter, Tamara A. Dailey, Harry A. Dailey, William N. Lanzilotta
Journal of Molecular Biology (23 October 2009) Volume 393(Issue 2) pp:308-319
Publication Date(Web):23 October 2009
DOI:10.1016/j.jmb.2009.08.042
Ferrochelatase (protoheme ferrolyase, E.C. 4.99.1.1) is the terminal enzyme in heme biosynthesis and catalyzes the insertion of ferrous iron into protoporphyrin IX to form protoheme IX (heme). Within the past two years, X-ray crystallographic data obtained with human ferrochelatase have clearly shown that significant structural changes occur during catalysis that are predicted to facilitate metal insertion and product release. One unanswered question about ferrochelatase involves defining the mechanism whereby some metals, such as divalent Fe, Co, Ni, and Zn, can be used by the enzyme in vitro to produce the corresponding metalloporphyrins, while other metals, such as divalent Mn, Hg, Cd, or Pb, are inhibitors of the enzyme. Through the use of high-resolution X-ray crystallography along with characterization of metal species via their anomalous diffraction, the identity and position of Hg, Cd, Ni, or Mn in the center of enzyme-bound porphyrin macrocycle were determined. When Pb, Hg, Cd, or Ni was present in the macrocycle, the conserved π helix was in the extended, partially unwound “product release” state. Interestingly, in the structure of ferrochelatase with Mn-porphyrin bound, the π helix is not extended or unwound and is in the “substrate-bound” conformation. These findings show that at least in the cases of Mn, Pb, Cd, and Hg, metal “inhibition” of ferrochelatase is not due to the inability of the enzyme to insert the metal into the macrocycle or by binding to a second metal binding site as has been previously proposed. Rather, inhibition occurs after metal insertion and results from poor or diminished product release. Possible explanations for the lack of product release are proposed herein.
Co-reporter:Amy E. Medlock, Tamara A. Dailey, Teresa A. Ross, Harry A. Dailey, William N. Lanzilotta
Journal of Molecular Biology (2 November 2007) Volume 373(Issue 4) pp:1006-1016
Publication Date(Web):2 November 2007
DOI:10.1016/j.jmb.2007.08.040
Ferrochelatase (protoheme ferrolyase, EC 4.99.1.1) is the terminal enzyme in heme biosynthesis and catalyzes the insertion of ferrous iron into protoporphyrin IX to form protoheme IX (heme). Due to the many critical roles of heme, synthesis of heme is required by the vast majority of organisms. Despite significant investigation of both the microbial and eukaryotic enzyme, details of metal chelation remain unidentified. Here we present the first structure of the wild-type human enzyme, a lead-inhibited intermediate of the wild-type enzyme with bound metallated porphyrin macrocycle, the product bound form of the enzyme, and a higher resolution model for the substrate-bound form of the E343K variant. These data paint a picture of an enzyme that undergoes significant changes in secondary structure during the catalytic cycle. The role that these structural alterations play in overall catalysis and potential protein–protein interactions with other proteins, as well as the possible molecular basis for these changes, is discussed. The atomic details and structural rearrangements presented herein significantly advance our understanding of the substrate binding mode of ferrochelatase and reveal new conformational changes in a structurally conserved π-helix that is predicted to have a central role in product release.
.jpg)
![Ferrate(2-), [3,7,12,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/1054400/18922-88-8.png)
![Ferrate(2-), [3,7,12,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/1054400/18922-88-8_b.png)


![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8.png)
![Ferrate(2-), [7,12-diethenyl-3,8,13,17-tetramethyl-21H,23H-porphine-2,18-dipropanoato(4-)-κN21,κN22,κN23,κN24]-, hydrogen (1:2), (SP-4-2)-](/data/chemimg/522800/14875-96-8_b.png)