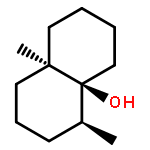
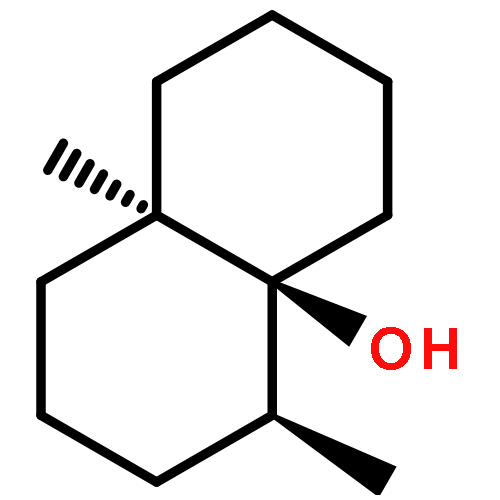
The biosynthesis of terpenes is catalysed by class I and II terpene cyclases. Here we present structural data from a class I hedycaryol synthase in complex with nerolidol, serving as a surrogate for the reaction intermediate nerolidyl diphosphate. This prefolded ligand allows mapping of the active site and hence the identification of a key carbonyl oxygen of Val179, a highly conserved helix break (G1/2) and its corresponding helix dipole. Stabilising the carbocation at the substrate's C1 position, these elements act in concert to catalyse the 1,10 ring closure, thereby exclusively generating the anti-Markovnikov product. The delineation of a general mechanistic scaffold was confirmed by site-specific mutations. This work serves as a basis for understanding carbocation chemistry in enzymatic reactions and should contribute to future application of these enzymes in organic synthesis.
A derivative of the pET28c(+) expression vector was constructed. It contains a yeast replication system (2μ origin of replication) and a yeast selectable marker (URA3), and can be used for gene cloning in yeast by efficient homologous recombination, and for heterologous expression in E. coli. The vector was used for the expression and chemical characterisation of three bacterial terpene cyclases.
We present crystallographic and functional data of selina-4(15),7(11)-diene synthase (SdS) from Streptomyces pristinaespiralis in its open and closed (ligand-bound) conformation. We could identify an induced-fit mechanism by elucidating a rearrangement of the G1/2 helix-break motif upon substrate binding. This rearrangement highlights a novel effector triad comprising the pyrophosphate sensor Arg178, the linker Asp181, and the effector Gly182-O. This structural motif is strictly conserved in class I terpene cyclases from bacteria, fungi, and plants, including epi-isozizaene synthase (3KB9), aristolochene synthase (4KUX), bornyl diphosphate synthase (1N20), limonene synthase (2ONG), 5-epi-aristolochene synthase (5EAT), and taxa-4(5),11(12)-diene synthase (3P5R). An elaborate structure-based mutagenesis in combination with analysis of the distinct product spectra confirmed the mechanistic models of carbocation formation and stabilization in SdS.
Volatiles emitted by the soil fungi Trichoderma viride 272 and Trichoderma asperellum 328 were collected by using the closed loop stripping analysis (CLSA) headspace technique, and the obtained extracts were analysed by GC/MS. Several alkyl- and alkenyl-2H-pyran-2-ones, including known compounds 6-pentyl-2H-pyran-2-one and (E)-6-(pent-1-en-1-yl)-2H-pyran-2-one, and the new derivatives (E)-6-(pent-2-en-1-yl)-2H-pyran-2-one, 6-propyl-2H-pyran-2-one, and 6-heptyl-2H-pyran-2-one were found. The alkenyl derivative (E)-6-(hept-1-en-1-yl)-2H-pyran-2-one, previously tentatively identified from a marine Botrytis by MS analysis, was also detected. All alkenyl pyrones were synthesised by using a reported Stille coupling followed by lactonisation, whereas the alkylated pyrones were obtained through a reported synthetic approach by radical bromination of 5-alkylpent-2-en-5-olides and dehydrobromination. Because the yields in both cases were not satisfactory and fell a long way short of the yields reported for similar compounds, all compounds were synthesised again using a gold-catalysed coupling of terminal alkynes with propiolic acid recently developed by Schreiber and co-workers, giving high yields in all cases. A comparison of the synthetic methods is given.
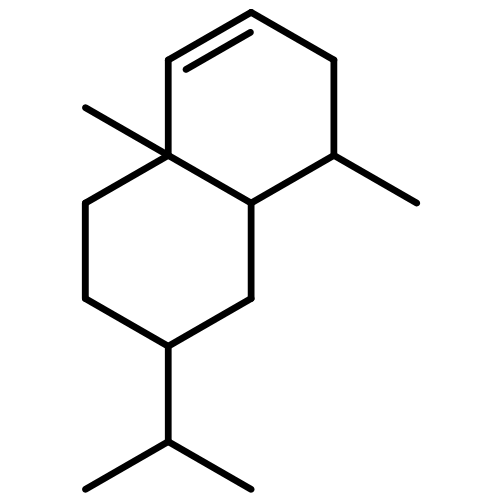
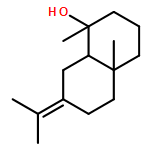
The volatile terpenes of 24 actinomycetes whose genomes have been sequenced (or are currently being sequenced) were collected by use of a closed-loop stripping apparatus and identified by GC/MS. The analytical data were compared against a phylogenetic analysis of all 192 currently available sequences of bacterial terpene cyclases (excluding geosmin and 2-methylisoborneol synthases). In addition to the several groups of terpenes with known biosynthetic origin, selinadienes were identified as a large group of biosynthetically related sesquiterpenes that are produced by several streptomycetes. The detection of a large number of previously unrecognised side products of known terpene cyclases proved to be particularly important for an in depth understanding of biosynthetic pathways to known terpenes in actinomycetes. Interpretation of the chemical analytical data in the context of the phylogenetic tree of bacterial terpene cyclases pointed to the function of three new enzymes: (E)-β-caryophyllene synthase, selina-3,7(11)-diene synthase and aristolochene synthase.
Several volatile lactones have been identified from the endophytic fungus Geniculosporium sp. isolated from the rockrose Cistus monspeliensis, commonly known as Montpelier cistus. The fungal volatiles were collected from agar-plate cultures by using a closed-loop stripping apparatus and the headspace extracts were analysed by GC–MS. Structures for these lactones were proposed from their mass spectral data. The suggested structures were verified by synthesis of reference compounds through a Diels–Alder/retro-Diels–Alder approach. This synthetic method was also successfully applied in the synthesis of butenolides that are signalling molecules from streptomycetes. For the enantioselective synthesis of these butenolides a modified route including an enantioselective Diels–Alder reaction was used.
The genomes of all bacteria with publicly available sequenced genomes have been screened for the presence of sesquiterpene cyclase homologues, resulting in the identification of 55 putative geosmin synthases, 23 homologues of 2-methylisoborneol synthases, and 98 other sesquiterpene cyclase homologues. Most of these enzymes by far were found in actinomycetes. The terpenoid volatiles from 35 strains, including 31 actinomycetes and four strains from other taxa, were collected by using a closed-loop stripping apparatus and identified by GC-MS. All of these bacteria apart from one strain encode sesquiterpene cyclase homologues in their genomes. The identified volatile terpenoids were grouped according to structural similarities and their biosynthetic relationship, and the results of these analyses were correlated to the available genome information, resulting in valuable new insights into bacterial terpene biosynthesis.
The volatiles released by several streptomycetes were collected by using a closed-loop stripping apparatus (CLSA) and analysed by GC–MS. The obtained headspace extracts of various species contained blastmycinone, a known degradation product of the fungicidal antibiotic, antimycin A3b, and several unknown derivatives. The suggested structures of these compounds, based on their mass spectra and GC retention indices, were confirmed by comparison to synthetic reference samples. Additional compounds found in the headspace extracts were butenolides formed from the blastmycinones by elimination of the carboxylic acid moiety. Analysis of a gene knockout mutant in the antimycin biosynthetic gene cluster demonstrated that all blastmycinones and butenolides are formed via the antimycin biosynthetic pathway. The structural variation of the blastmycinones identified here is much larger than within the known antimycins, thus suggesting that several antimycin derivatives remain to be discovered.
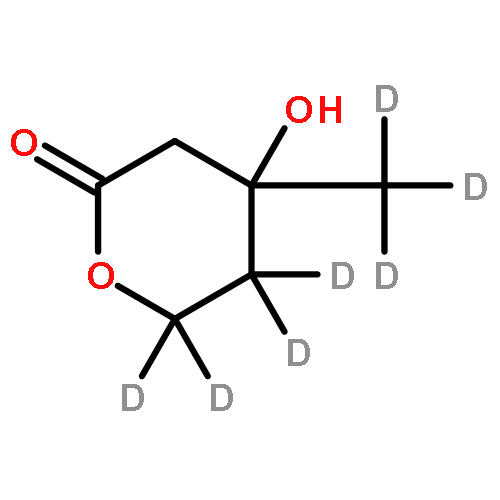
A synthetic route was developed for the preparation of deuterated mevalonolactones. Using low-cost deuterated reagents, this route allows for the independent introduction of deuterium labeling into any carbon position or into any combination of positions. Following this approach, the synthesis of [6,6,6-2H3]mevalonolactone, [4,4,6,6,6-2H5]mevalonolactone, [5,5-2H2]mevalonolactone, [5,5,6,6,6-2H5]mevalonolactone, and [2,2,6,6,6-2H5]mevalonolactone is described.
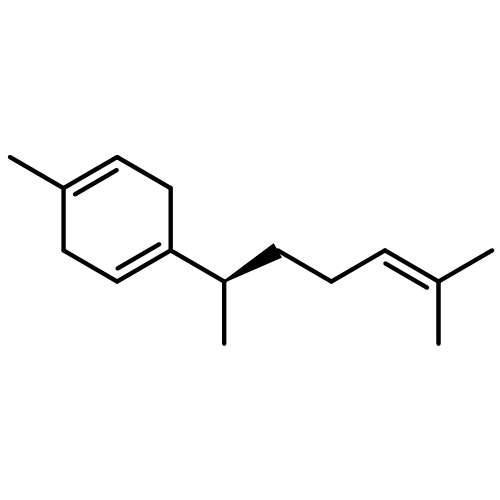
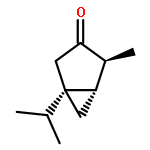
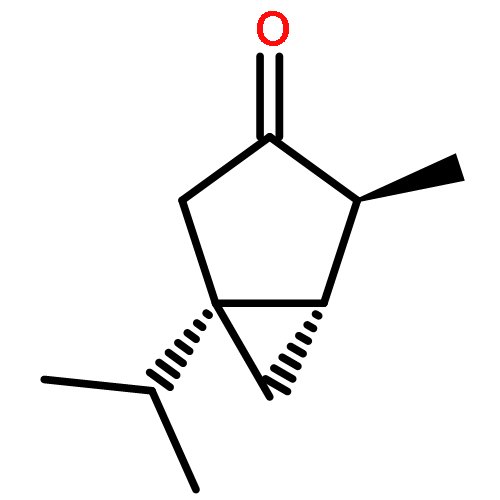
The volatiles released by several strains of Trichoderma fungi were collected using a closed-loop stripping apparatus and analysed by GC–MS. Most of the investigated strains released the two structurally related spirocyclic sesquiterpenoids trichoacorenol, previously identified in Trichoderma koningii (Huang et al., 1995), and acorenone. A new enantioselective synthesis of the enantiomer of the natural spirocyclic sesquiterpene acorenone and the first enantioselective synthesis of the related compound ent-trichoacorenol have been completed by a chiral-pool approach starting from (+)-(R)-pulegone using ring-closing metathesis as the key step. The absolute configuration of natural trichoacorenol from Trichoderma harzianum sp. 714 has been assigned by chiral GC–MS analysis and is the same as that in T. koningii.
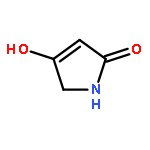
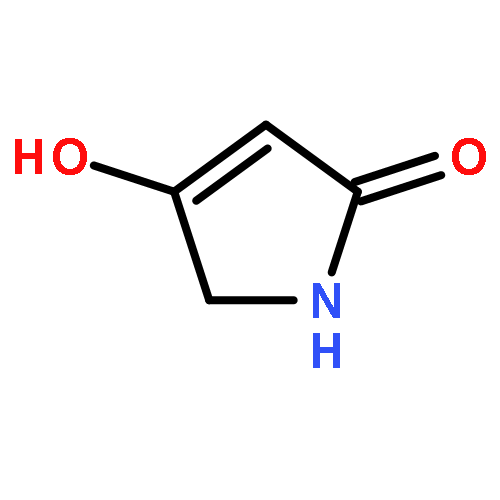
The total syntheses of the structurally unique and moderately cytotoxic pyrrolosesquiterpenoid glaciapyrrol A that has been isolated from a marine streptomycete by Macherla et al.1 and of seven of its stereoisomers have been performed from geraniol or nerol, respectively, using a known diastereoselective Ru-catalysed approach for the synthesis of tetrahydrofurans previously reported by Stark and co-workers.5 Comparison of 1H and 13C NMR data unambiguously clarified the relative configuration of natural glaciapyrrol A that was previously only partly solved from the available NMR data. An enantioselective synthesis was carried out resulting in the unnatural enantiomer (11S,12R,15R)-(−)-glaciapyrrol A. These data establish the absolute configuration of the natural product as (11R,12S,15S)-(+)-glaciapyrrol A.
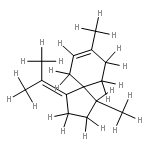
The volatiles of the fungus Fusarium verticillioides were analysed by GC-MS. Sesquiterpenes dominated, with trichodiene as the principle component. Several other sesquiterpenes were detected in low amounts that were unambiguously identified from their mass spectra and retention indices. The absolute configurations of (R)-β-bisabolene, (R)-cuparene, (+)-β-barbatene, (−)-α-cedrene, (+)-β-cedrene, and (+)-α-funebrene originating from different key cationic intermediates, were determined by chiral GC-MS and proved to be related to the trichodiene stereostructure. The unusual compound (E)-iso-γ-bisabolene was also found corroborating a previously suggested mechanism for the cyclisation of the bisabolyl to the cuprenyl cation that is based on quantum mechanical calculations (Y. J. Hong, D. J. Tantillo, Org. Lett.2006, 8, 4601–4604). These analyses resulted in a revised biosynthesis scheme to trichodiene and the side products of the responsible terpene cyclase, trichodiene synthase, an enzyme that is well characterised from Fusarium sporotrichioides. Feeding studies with several deuterated mevalonolactone isotopomers unravelled stereochemical aspects of the late cyclisations towards trichodiene.
The fungus Fusarium fujikuroi IMI58289 emits a complex pattern of volatile terpenoids including two major compounds, the sesquiterpene alcohol α-acorenol and the diterpene ent-kaurene. ent-Kaurene is the precursor for the phytohormone gibberellic acid (GA3) and is produced from geranylgeranyl diphosphate (GGPP) via ent-copalyl diphosphate by the bifunctional ent-copalyl diphosphate/ent-kaurene synthase (CPS/KS). Several structurally related diterpenes were identified as side products of the CPS/KS. Deletion of the cps/ks gene or the whole GA3 biosynthetic gene cluster resulted in completely abolished diterpene production. Mutants with deletions of the cytochrome P450 monooxygenase gene P450-4, which is responsible for the three oxidation steps from ent-kaurene to ent-kaurenoic acid en route to GA3, accumulate diterpene hydrocarbons. Feeding with [6,6,6-2H3] mevalonolactone gave insights into the stereochemistry of the GGPP cyclisation, which operates with a chair–chair-“antipodal” fold. A rational biosynthetic scheme for all identified sesquiterpenes demonstrated their formation from farnesyl diphosphate (FPP) via three alternative initial cyclisations. Genome sequencing revealed the presence of five putative sesquiterpene synthase genes in the F. fujikuroi genome. The structures of several trace compounds from other classes have been identified as new natural products; these were delineated from their mass spectra and unambiguously assigned by comparison to synthetic references.
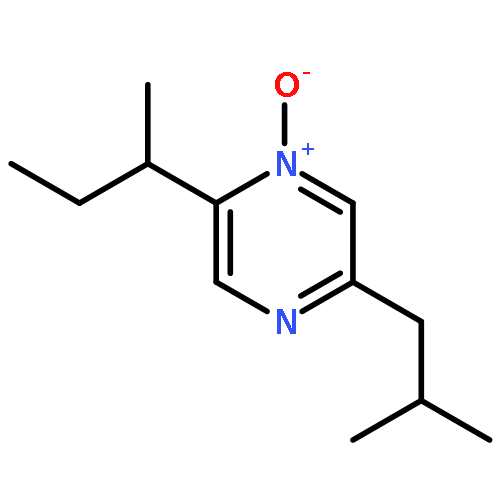
The volatile compounds released by Corynebacterium glutamicum were collected by use of the CLSA technique (closed-loop stripping apparatus) and analysed by GC-MS. The headspace extracts contained several acyloins and pyrazines that were identified by their synthesis or comparison to commercial standards. Feeding experiments with [2H7]acetoin resulted in the incorporation of labelling into trimethylpyrazine and tetramethylpyrazine. Several deletion mutants targeting genes of the primary metabolism were constructed to elucidate the biosynthetic pathway to pyrazines in detail. A deletion mutant of the ketol-acid reductoisomerase was not able to convert the acetoin precursor (S)-2-acetolactate into the pathway intermediate (R)-2,3-dihydroxy-3-methylbutanoate to the branched amino acids. This mutant requires valine, leucine, and isoleucine for growth and produces significantly higher amounts and more different compounds of the acyloin and pyrazine classes. Gene deletion of the acetolactate synthase (AS) resulted in a mutant that is not able to convert pyruvate into (S)-2-acetolactate. This mutant also requires branched amino acids and produces only very small amounts of pyrazines likely from valine via the valine biosynthetic pathway operating in reverse order. A ΔASΔKR double mutant was constructed that does not produce any pyrazines at all. These results open up a detailed biosynthetic model for the formation of alkylated pyrazines via acyloins.
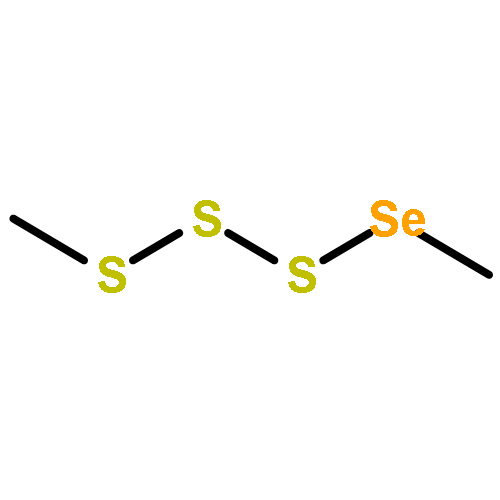
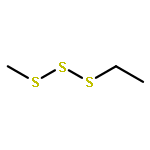
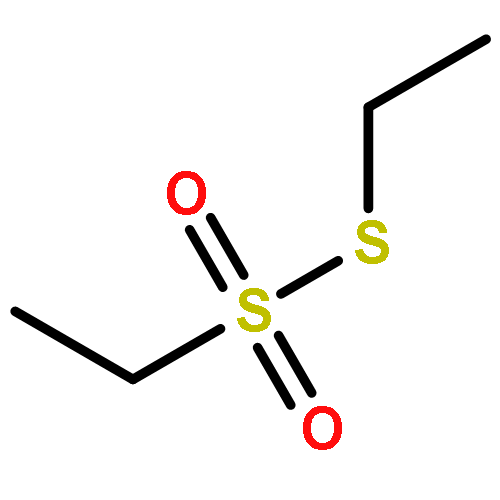
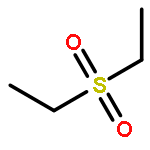
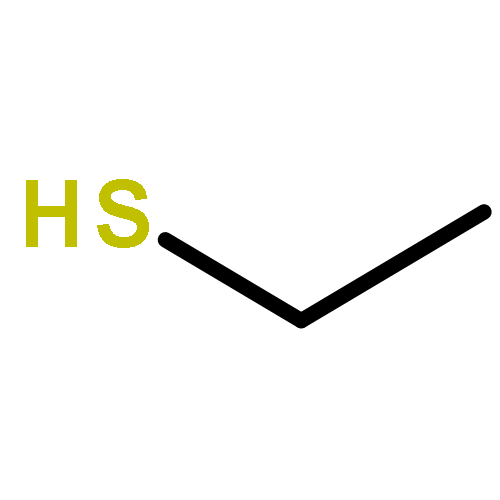
The volatiles released by Phaeobacter gallaeciensis, Oceanibulbus indolifex and Dinoroseobacter shibae have been investigated by GC-MS, and several MeSH-derived sulfur volatiles have been identified. An important sulfur source in the oceans is the algal metabolite dimethylsulfoniopropionate (DMSP). Labelled [2H6]DMSP was fed to the bacteria to investigate the production of volatiles from this compound through the lysis pathway to [2H6]dimethylsulfide or the demethylation pathway to [2H3]-3-(methylmercapto)propionic acid and lysis to [2H3]MeSH. [2H6]DMSP was efficiently converted to [2H3]MeSH by all three species. Several DMSP derivatives were synthesised and used in feeding experiments. Strong dealkylation activity was observed for the methylated ethyl methyl sulfoniopropionate and dimethylseleniopropionate, as indicated by the formation of EtSH- and MeSeH-derived volatiles, whereas no volatiles were formed from dimethyltelluriopropionate. In contrast, the dealkylation activity for diethylsulfoniopropionate was strongly reduced, resulting in only small amounts of EtSH-derived volatiles accompanied by diethyl sulfide in P. gallaeciensis and O. indolifex, while D. shibae produced the related oxidation product diethyl sulfone. The formation of diethyl sulfide and diethyl sulfone requires the lysis pathway, which is not active for [2H6]DMSP. These observations can be explained by a shifted distribution between the two competing pathways due to a blocked dealkylation of ethylated substrates.








![2-OCTANONE, 8-[(TETRAHYDRO-2H-PYRAN-2-YL)OXY]-](http://img.cochemist.com/ccimg/64900/64841-41-4.png)
![2-OCTANONE, 8-[(TETRAHYDRO-2H-PYRAN-2-YL)OXY]-](http://img.cochemist.com/ccimg/64900/64841-41-4_b.png)

























































































































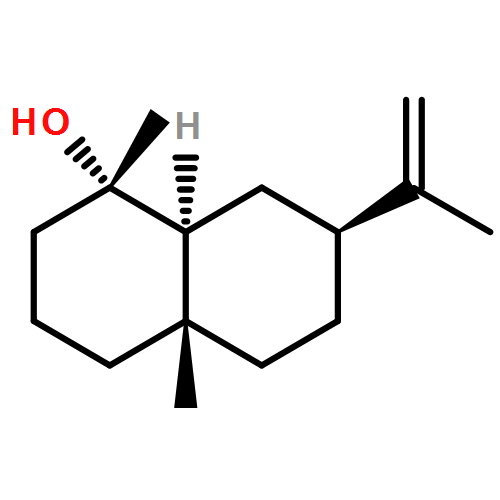








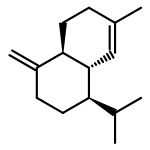
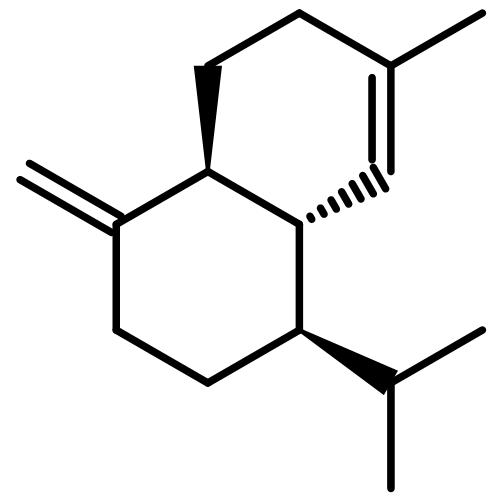






![2-ETHYL-1,6-DIOXASPIRO[4.4]-NONANE](http://img.cochemist.com/ccimg/38500/38401-84-2.png)
![2-ETHYL-1,6-DIOXASPIRO[4.4]-NONANE](http://img.cochemist.com/ccimg/38500/38401-84-2_b.png)


![(-)-(1R,4S,5R,6R,7S,10R)-7-isopropyl-4,10-dimethyltricyclo[4.4.0.0(1,5)]decan-4-ol](http://img.cochemist.com/ccimg/38300/38230-60-3.png)
![(-)-(1R,4S,5R,6R,7S,10R)-7-isopropyl-4,10-dimethyltricyclo[4.4.0.0(1,5)]decan-4-ol](http://img.cochemist.com/ccimg/38300/38230-60-3_b.png)
















































![Butanoic acid,2-methyl-,(2R,3S,6S,7R,8R)-3-[[3-(formylamino)-2-hydroxybenzoyl]amino]-8-hexyl-2,6-dimethyl-4,9-dioxo-1,5-dioxonan-7-ylester](http://img.cochemist.com/ccimg/28100/28068-16-8.png)
![Butanoic acid,2-methyl-,(2R,3S,6S,7R,8R)-3-[[3-(formylamino)-2-hydroxybenzoyl]amino]-8-hexyl-2,6-dimethyl-4,9-dioxo-1,5-dioxonan-7-ylester](http://img.cochemist.com/ccimg/28100/28068-16-8_b.png)
![Bicyclo[2.2.1]heptane,1,7,7-trimethyl-2-methylene-](http://img.cochemist.com/ccimg/27600/27538-47-2.png)
![Bicyclo[2.2.1]heptane,1,7,7-trimethyl-2-methylene-](http://img.cochemist.com/ccimg/27600/27538-47-2_b.png)






![4-[(1E)-1,5-dimethylhexa-1,4-dien-1-yl]-1-methylcyclohexene](http://img.cochemist.com/ccimg/25600/25532-79-0.png)
![4-[(1E)-1,5-dimethylhexa-1,4-dien-1-yl]-1-methylcyclohexene](http://img.cochemist.com/ccimg/25600/25532-79-0_b.png)












![6-[(1E,3E,5E)-hepta-1,3,5-trien-1-yl]-2,3,4,5-tetrahydropyridine](http://img.cochemist.com/ccimg/24000/23943-03-5.png)
![6-[(1E,3E,5E)-hepta-1,3,5-trien-1-yl]-2,3,4,5-tetrahydropyridine](http://img.cochemist.com/ccimg/24000/23943-03-5_b.png)
![1H-Cyclopenta[1,3]cyclopropa[1,2]benzen-3-ol,octahydro-3,7-dimethyl-4-(1-methylethyl)-, (3S,3aR,3bR,4S,7R,7aR)-](http://img.cochemist.com/ccimg/23500/23445-02-5.png)
![1H-Cyclopenta[1,3]cyclopropa[1,2]benzen-3-ol,octahydro-3,7-dimethyl-4-(1-methylethyl)-, (3S,3aR,3bR,4S,7R,7aR)-](http://img.cochemist.com/ccimg/23500/23445-02-5_b.png)
![3-Buten-2-one,4-(2,2,6-trimethyl-7-oxabicyclo[4.1.0]hept-1-yl)-](http://img.cochemist.com/ccimg/23300/23267-57-4.png)
![3-Buten-2-one,4-(2,2,6-trimethyl-7-oxabicyclo[4.1.0]hept-1-yl)-](http://img.cochemist.com/ccimg/23300/23267-57-4_b.png)




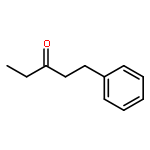
![Benzene, [(3-butynyloxy)methyl]-](http://img.cochemist.com/ccimg/22300/22273-77-4.png)
![Benzene, [(3-butynyloxy)methyl]-](http://img.cochemist.com/ccimg/22300/22273-77-4_b.png)










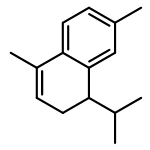
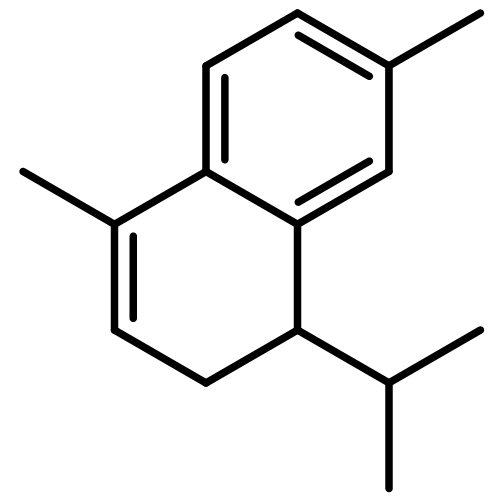
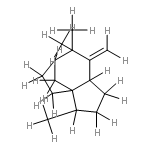
![2-[(2R,8R,8aS)-8,8a-dimethyl-1,2,3,4,6,7,8,8a-octahydronaphthalen-2-yl]propan-2-ol](http://img.cochemist.com/ccimg/20500/20489-45-6.png)
![2-[(2R,8R,8aS)-8,8a-dimethyl-1,2,3,4,6,7,8,8a-octahydronaphthalen-2-yl]propan-2-ol](http://img.cochemist.com/ccimg/20500/20489-45-6_b.png)







![(3R,6S,6aR,9S)-2,2,6,9-tetramethyloctahydro-2H-3,9a-methanocyclopenta[b]oxocine](http://img.cochemist.com/ccimg/20200/20149-50-2.png)
![(3R,6S,6aR,9S)-2,2,6,9-tetramethyloctahydro-2H-3,9a-methanocyclopenta[b]oxocine](http://img.cochemist.com/ccimg/20200/20149-50-2_b.png)






![Spiro[5.5]undeca-1,8-diene,1,5,5,9-tetramethyl-, (6R)-](http://img.cochemist.com/ccimg/20000/19912-83-5.png)
![Spiro[5.5]undeca-1,8-diene,1,5,5,9-tetramethyl-, (6R)-](http://img.cochemist.com/ccimg/20000/19912-83-5_b.png)











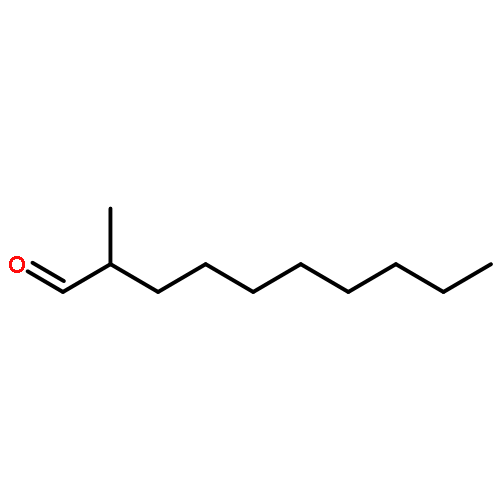






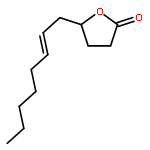
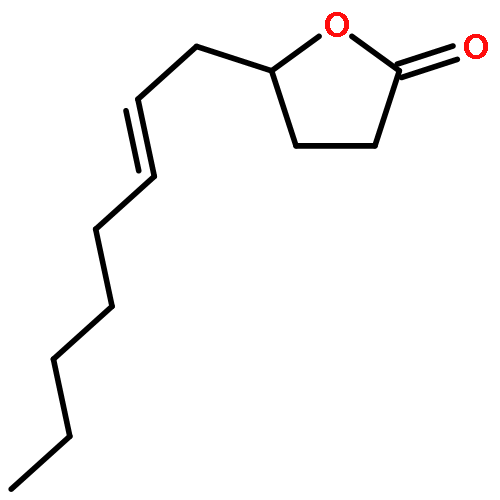




![Bicyclo[3.1.1]hept-2-ene, 2,6-dimethyl-6-(4-methyl-3-penten-1-yl)-, (1R,5R,6R)-rel-](/data/chemimg/1767200/18252-46-5.png)
![Bicyclo[3.1.1]hept-2-ene, 2,6-dimethyl-6-(4-methyl-3-penten-1-yl)-, (1R,5R,6R)-rel-](/data/chemimg/1767200/18252-46-5_b.png)

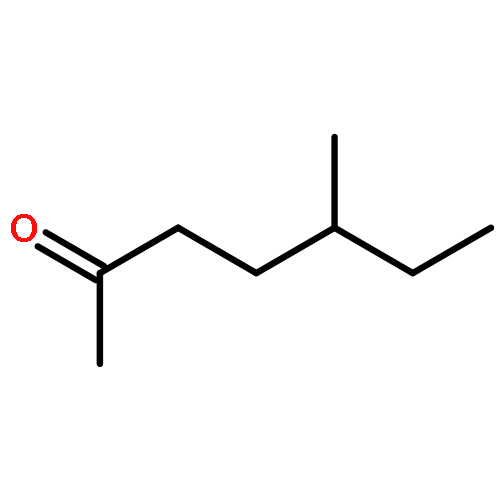






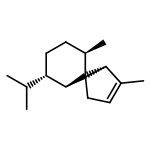
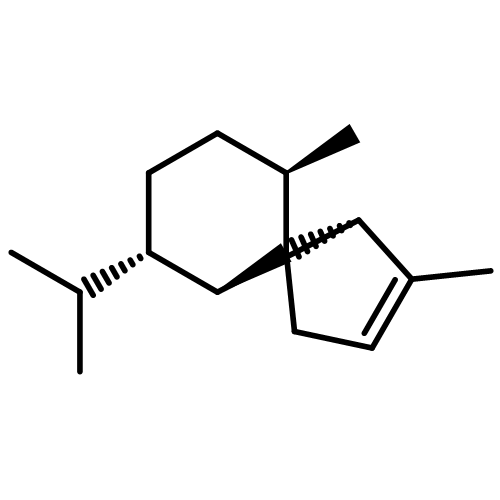




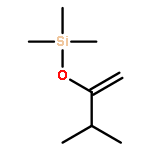

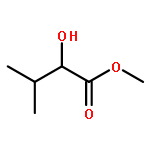
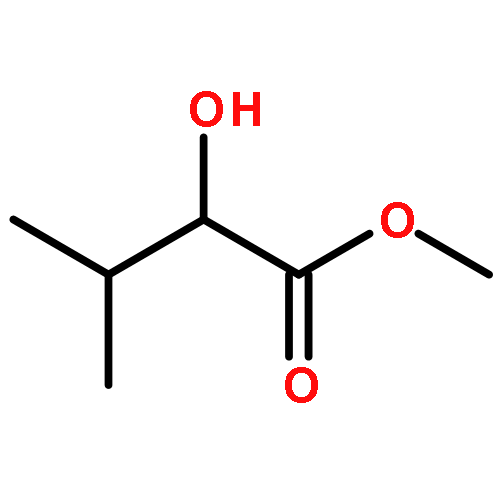
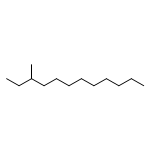
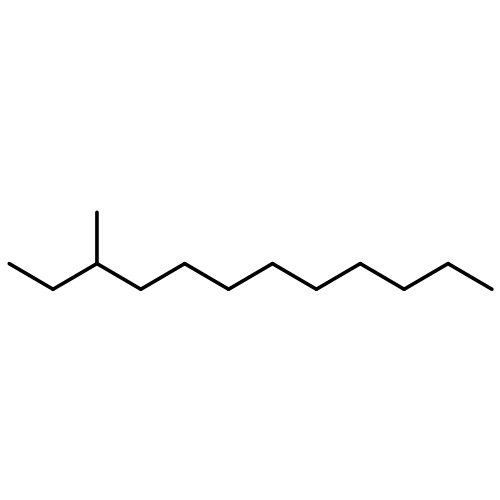










![(1α,2β,5α)-2-methyl-5-(1-methylethyl)bicyclo[3.1.0]hexan-2-ol](http://img.cochemist.com/ccimg/15600/15537-55-0.png)
![(1α,2β,5α)-2-methyl-5-(1-methylethyl)bicyclo[3.1.0]hexan-2-ol](http://img.cochemist.com/ccimg/15600/15537-55-0_b.png)
![Bicyclo[3.1.1]heptane, 6-methyl-2-methylene-6-(4-methyl-3-pentenyl)-,(1S,5S,6R)-](http://img.cochemist.com/ccimg/15500/15438-94-5.png)
![Bicyclo[3.1.1]heptane, 6-methyl-2-methylene-6-(4-methyl-3-pentenyl)-,(1S,5S,6R)-](http://img.cochemist.com/ccimg/15500/15438-94-5_b.png)
![Spiro[5.5]undec-2-ene, 3,7,7-trimethyl-11-methylene-](http://img.cochemist.com/ccimg/15500/15401-86-2.png)
![Spiro[5.5]undec-2-ene, 3,7,7-trimethyl-11-methylene-](http://img.cochemist.com/ccimg/15500/15401-86-2_b.png)



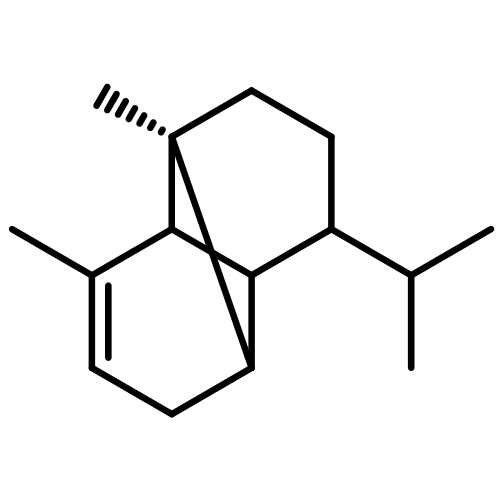








![(1R,4R,6S)-1-Methyl-4-(prop-1-en-2-yl)-7-oxabicyclo[4.1.0]heptane](http://img.cochemist.com/ccimg/13900/13837-75-7.png)
![(1R,4R,6S)-1-Methyl-4-(prop-1-en-2-yl)-7-oxabicyclo[4.1.0]heptane](http://img.cochemist.com/ccimg/13900/13837-75-7_b.png)




![Bicyclo[2.2.1]heptane, 1,3,3-trimethyl-2-methylene-](http://img.cochemist.com/ccimg/13600/13567-57-2.png)
![Bicyclo[2.2.1]heptane, 1,3,3-trimethyl-2-methylene-](http://img.cochemist.com/ccimg/13600/13567-57-2_b.png)


![(1S,5S)-4,6-DIMETHYL-6-(4-METHYLPENT-3-ENYL)BICYCLO[3.1.1]HEPT-3-ENE](http://img.cochemist.com/ccimg/13500/13474-59-4.png)
![(1S,5S)-4,6-DIMETHYL-6-(4-METHYLPENT-3-ENYL)BICYCLO[3.1.1]HEPT-3-ENE](http://img.cochemist.com/ccimg/13500/13474-59-4_b.png)









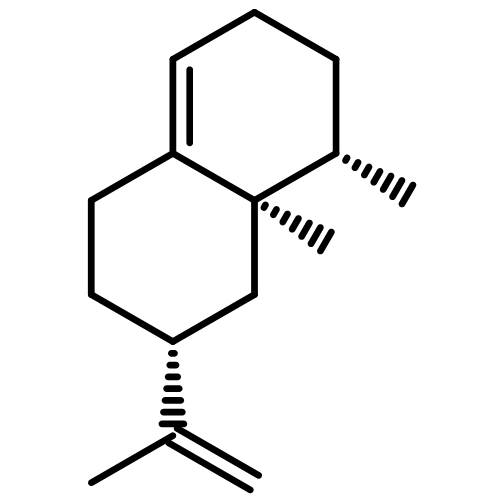




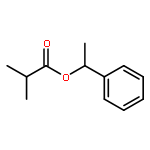
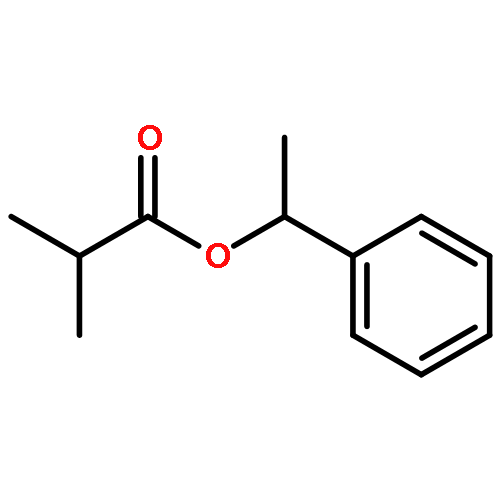












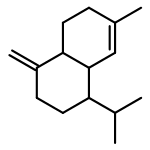



![Diphosphoric acid,P-[(2E,6E,10E)-3,7,11,15-tetramethyl-2,6,10,14-hexadecatetraen-1-yl] ester](http://img.cochemist.com/ccimg/6700/6699-20-3.png)
![Diphosphoric acid,P-[(2E,6E,10E)-3,7,11,15-tetramethyl-2,6,10,14-hexadecatetraen-1-yl] ester](http://img.cochemist.com/ccimg/6700/6699-20-3_b.png)




![2-morpholin-4-yl-N-(4-{5-[(morpholin-4-ylacetyl)amino]-1,3-benzoxazol-2-yl}phenyl)acetamide](http://img.cochemist.com/ccimg/6200/6158-93-6.png)
![2-morpholin-4-yl-N-(4-{5-[(morpholin-4-ylacetyl)amino]-1,3-benzoxazol-2-yl}phenyl)acetamide](http://img.cochemist.com/ccimg/6200/6158-93-6_b.png)


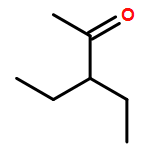

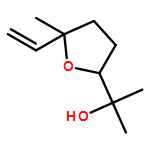
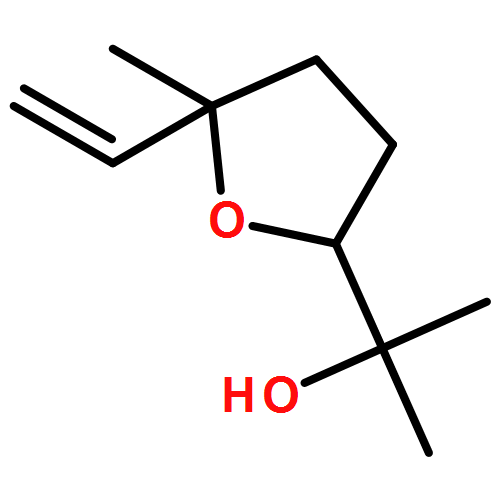





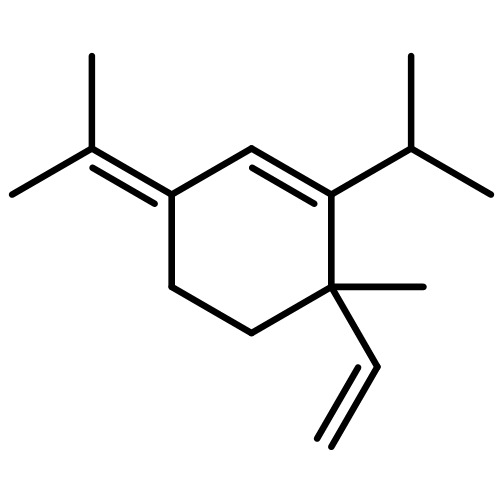
![2-[(4-methoxyphenyl)amino]-4,6-dimethylpyridine-3-carboxamide](http://img.cochemist.com/ccimg/6000/5947-50-2.png)
![2-[(4-methoxyphenyl)amino]-4,6-dimethylpyridine-3-carboxamide](http://img.cochemist.com/ccimg/6000/5947-50-2_b.png)


































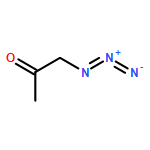
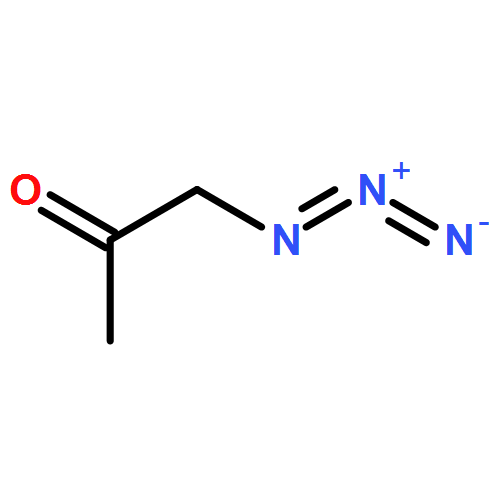


![1-[(1S)-1,5-dimethylhex-4-en-1-yl]-4-methylbenzene](http://img.cochemist.com/ccimg/4200/4176-06-1.png)
![1-[(1S)-1,5-dimethylhex-4-en-1-yl]-4-methylbenzene](http://img.cochemist.com/ccimg/4200/4176-06-1_b.png)
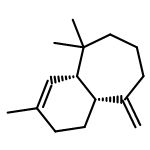
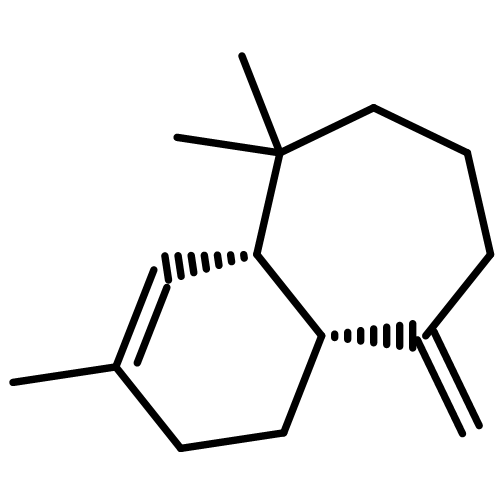
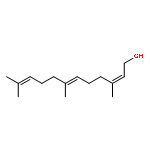
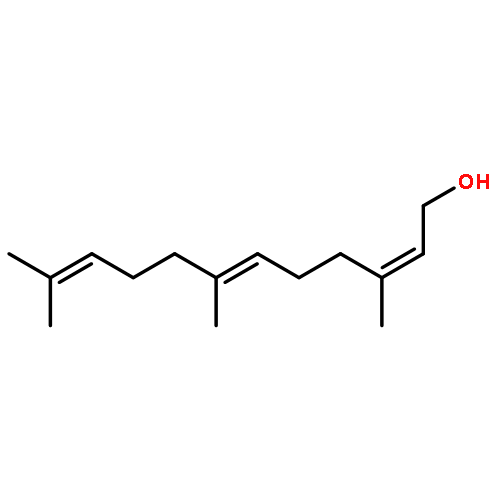







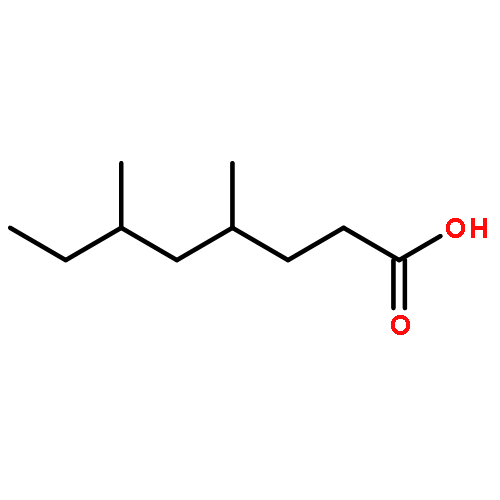








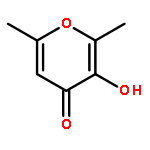







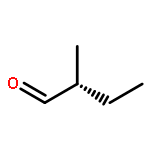
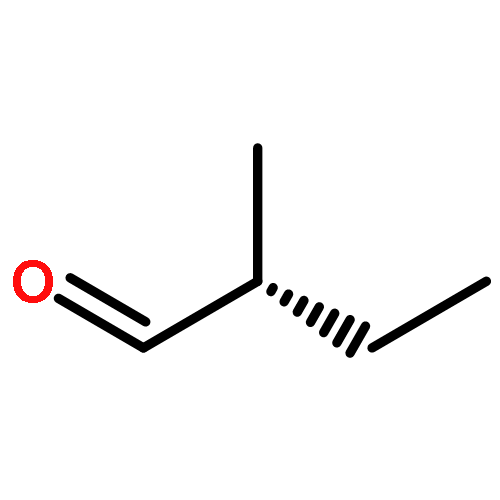






![2-Propanone, 1-[[(4-methylphenyl)sulfonyl]oxy]-](http://img.cochemist.com/ccimg/1700/1666-19-9.png)
![2-Propanone, 1-[[(4-methylphenyl)sulfonyl]oxy]-](http://img.cochemist.com/ccimg/1700/1666-19-9_b.png)




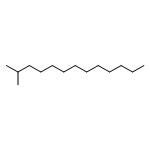
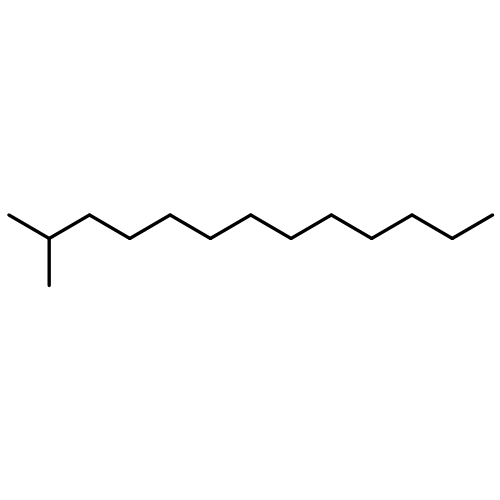








![1H-Naphtho[2,1-b]pyran,3-ethenyldodecahydro-3,4a,7,7,10a-pentamethyl-, (3S,4aR,6aS,10aS,10bR)-](http://img.cochemist.com/ccimg/1300/1227-93-6.png)
![1H-Naphtho[2,1-b]pyran,3-ethenyldodecahydro-3,4a,7,7,10a-pentamethyl-, (3S,4aR,6aS,10aS,10bR)-](http://img.cochemist.com/ccimg/1300/1227-93-6_b.png)



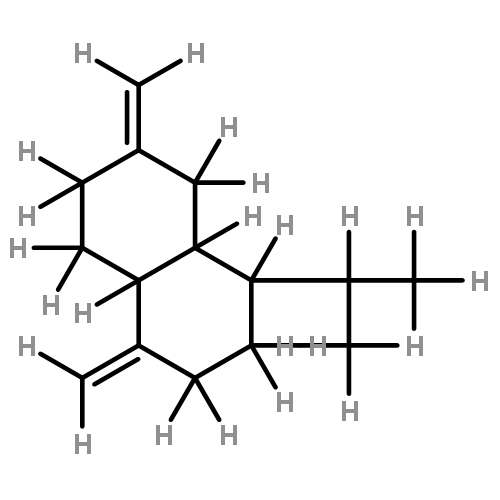






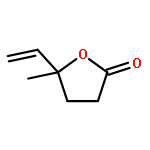


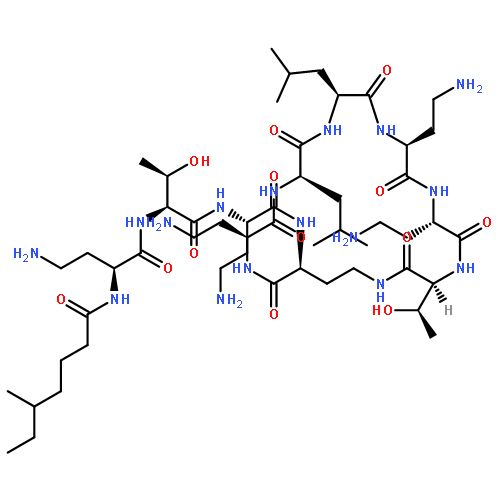
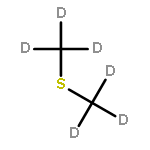
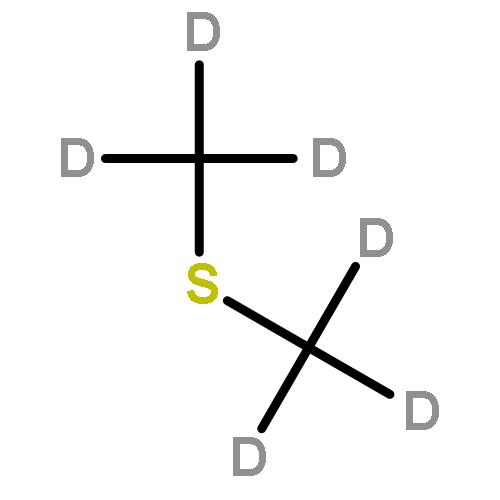

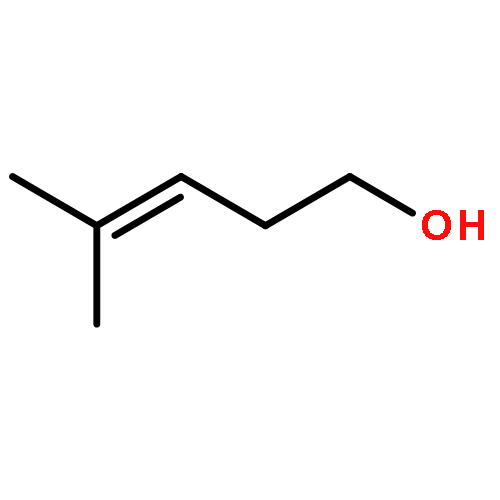


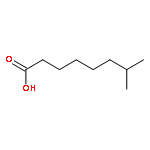


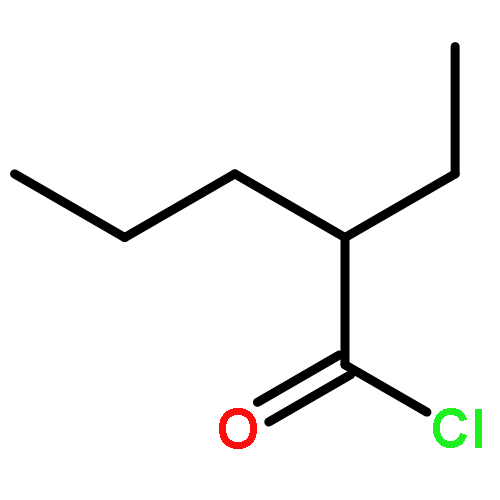

![Butanoic acid, 2(or3)-methyl-,(2R,3S,6S,7R,8R)-3-[[3-(formylamino)-2-hydroxybenzoyl]amino]-8-hexyl-2,6-dimethyl-4,9-dioxo-1,5-dioxonan-7-ylester (9CI)](http://img.cochemist.com/ccimg/700/642-15-9.png)
![Butanoic acid, 2(or3)-methyl-,(2R,3S,6S,7R,8R)-3-[[3-(formylamino)-2-hydroxybenzoyl]amino]-8-hexyl-2,6-dimethyl-4,9-dioxo-1,5-dioxonan-7-ylester (9CI)](http://img.cochemist.com/ccimg/700/642-15-9_b.png)






![1H-Naphtho[2,1-b]pyran,3-ethenyldodecahydro-3,4a,7,7,10a-pentamethyl-, (3R,4aR,6aS,10aS,10bR)- (9CI)](http://img.cochemist.com/ccimg/600/596-84-9.png)
![1H-Naphtho[2,1-b]pyran,3-ethenyldodecahydro-3,4a,7,7,10a-pentamethyl-, (3R,4aR,6aS,10aS,10bR)- (9CI)](http://img.cochemist.com/ccimg/600/596-84-9_b.png)









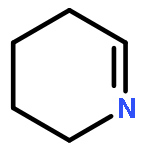
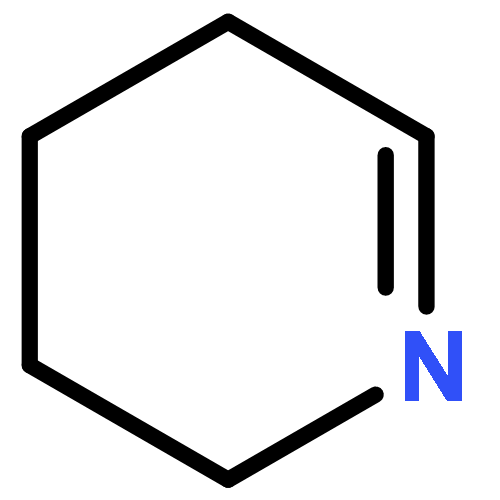





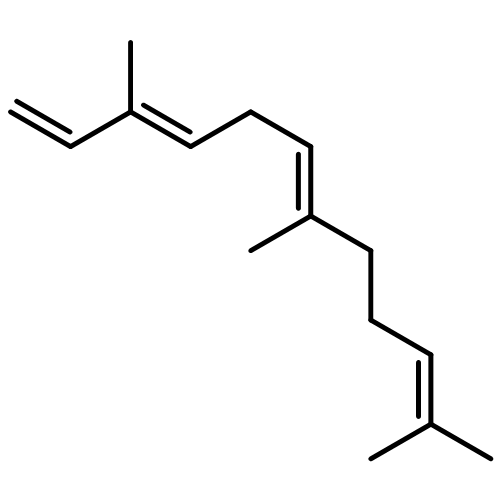
![1H-Cycloprop[e]azulene,1a,2,3,4,4a,5,6,7b-octahydro-1,1,4,7-tetramethyl-, (1aR,4R,4aR,7bS)-](http://img.cochemist.com/ccimg/500/489-40-7.png)
![1H-Cycloprop[e]azulene,1a,2,3,4,4a,5,6,7b-octahydro-1,1,4,7-tetramethyl-, (1aR,4R,4aR,7bS)-](http://img.cochemist.com/ccimg/500/489-40-7_b.png)




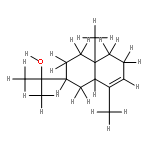
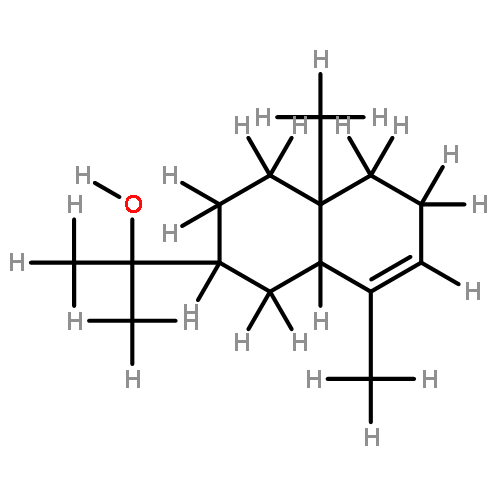
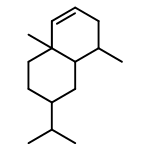



![Cyclopropa[d]naphthalene,1,1a,4,4a,5,6,7,8-octahydro-2,4a,8,8-tetramethyl-, (1aS,4aS,8aS)-](http://img.cochemist.com/ccimg/500/470-40-6.png)
![Cyclopropa[d]naphthalene,1,1a,4,4a,5,6,7,8-octahydro-2,4a,8,8-tetramethyl-, (1aS,4aS,8aS)-](http://img.cochemist.com/ccimg/500/470-40-6_b.png)




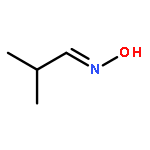
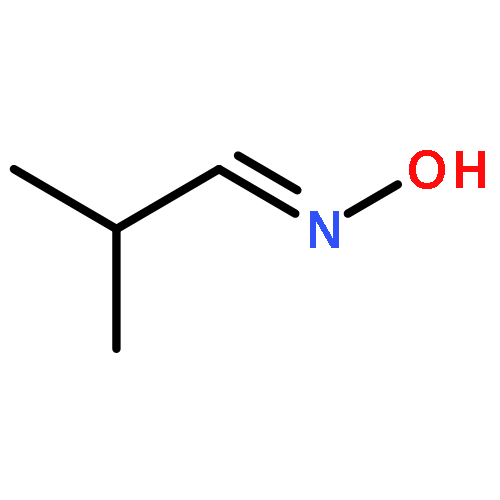


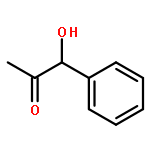
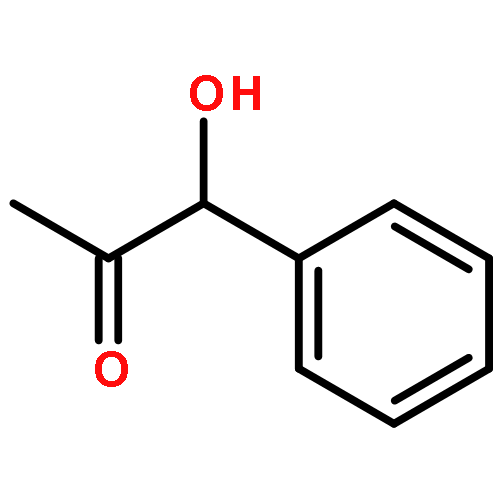

![1,7,7-TRIMETHYLBICYCLO[2.2.1]HEPTAN-2-OL ACETATE](http://img.cochemist.com/ccimg/100/76-49-3.png)
![1,7,7-TRIMETHYLBICYCLO[2.2.1]HEPTAN-2-OL ACETATE](http://img.cochemist.com/ccimg/100/76-49-3_b.png)













![Spiro[4.5]dec-7-ene, 1,8-dimethyl-4-(1-methylethenyl)-, (1R,4R,5S)-](http://img.cochemist.com/ccimg/729700/729602-94-2.png)
![Spiro[4.5]dec-7-ene, 1,8-dimethyl-4-(1-methylethenyl)-, (1R,4R,5S)-](http://img.cochemist.com/ccimg/729700/729602-94-2_b.png)

























![3,4,7,7-TETRAMETHYLBICYCLO[2.2.1]HEPT-2-ENE](http://img.cochemist.com/ccimg/72600/72540-93-3.png)
![3,4,7,7-TETRAMETHYLBICYCLO[2.2.1]HEPT-2-ENE](http://img.cochemist.com/ccimg/72600/72540-93-3_b.png)
![Cyclohexene, 3-methylene-6-[(1S)-1,2,2-trimethylcyclopentyl]-, (6R)-](http://img.cochemist.com/ccimg/66400/66389-22-8.png)
![Cyclohexene, 3-methylene-6-[(1S)-1,2,2-trimethylcyclopentyl]-, (6R)-](http://img.cochemist.com/ccimg/66400/66389-22-8_b.png)







![(1aR,2R,3R,3'S,3aR,5R,7bS)-3'-formyl-3,3',3a-trimethyl-6-oxo-2,3,3a,4,6,7b-hexahydro-1aH-spiro[naphtho[1,2-b]oxirene-5,2'-oxiran]-2-yl acetate](http://img.cochemist.com/ccimg/56300/56299-00-4.png)
![(1aR,2R,3R,3'S,3aR,5R,7bS)-3'-formyl-3,3',3a-trimethyl-6-oxo-2,3,3a,4,6,7b-hexahydro-1aH-spiro[naphtho[1,2-b]oxirene-5,2'-oxiran]-2-yl acetate](http://img.cochemist.com/ccimg/56300/56299-00-4_b.png)





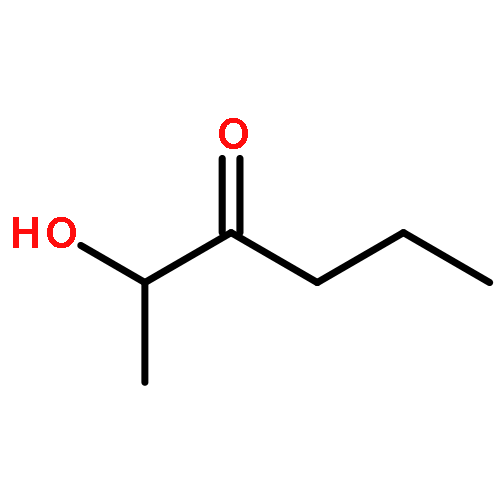












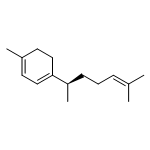

![Cyclohexene,1,4-dimethyl-4-[(1S)-1-methyl-2-methylenecyclopentyl]-, (4S)-](http://img.cochemist.com/ccimg/28700/28624-60-4.png)
![Cyclohexene,1,4-dimethyl-4-[(1S)-1-methyl-2-methylenecyclopentyl]-, (4S)-](http://img.cochemist.com/ccimg/28700/28624-60-4_b.png)
![Spiro[4.5]dec-7-ene, 1,8-dimethyl-4-(1-methylethenyl)-, (1R,4S,5R)-](http://img.cochemist.com/ccimg/28500/28477-64-7.png)
![Spiro[4.5]dec-7-ene, 1,8-dimethyl-4-(1-methylethenyl)-, (1R,4S,5R)-](http://img.cochemist.com/ccimg/28500/28477-64-7_b.png)
![Spiro[4.5]dec-7-ene, 1,8-dimethyl-4-(1-methylethylidene)-, (1R,5S)-](http://img.cochemist.com/ccimg/28500/28400-12-6.png)
![Spiro[4.5]dec-7-ene, 1,8-dimethyl-4-(1-methylethylidene)-, (1R,5S)-](http://img.cochemist.com/ccimg/28500/28400-12-6_b.png)



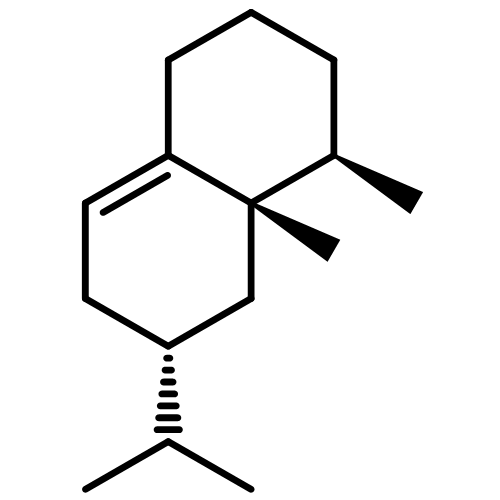

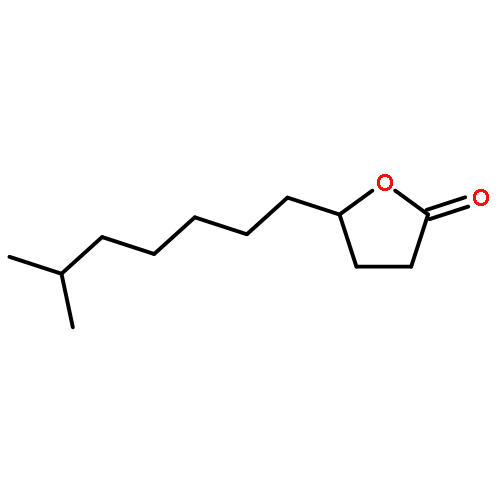


![Spiro[4.5]dec-7-ene,1,8-dimethyl-4-(1-methylethenyl)-, (1R,4S,5S)-](http://img.cochemist.com/ccimg/24100/24048-44-0.png)
![Spiro[4.5]dec-7-ene,1,8-dimethyl-4-(1-methylethenyl)-, (1R,4S,5S)-](http://img.cochemist.com/ccimg/24100/24048-44-0_b.png)











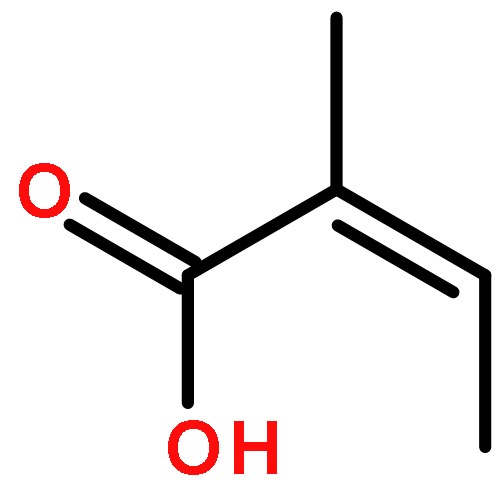






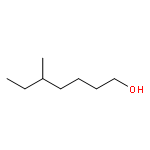
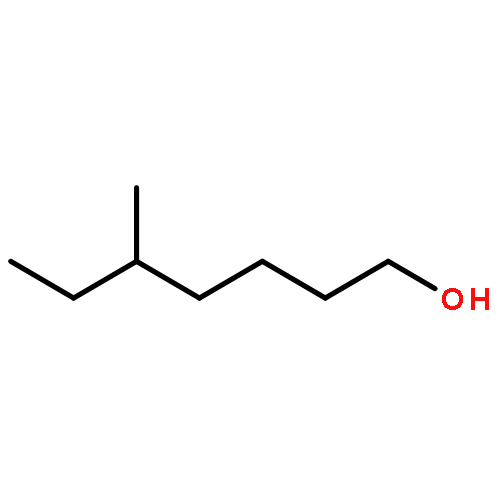




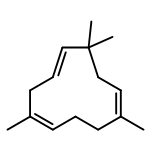
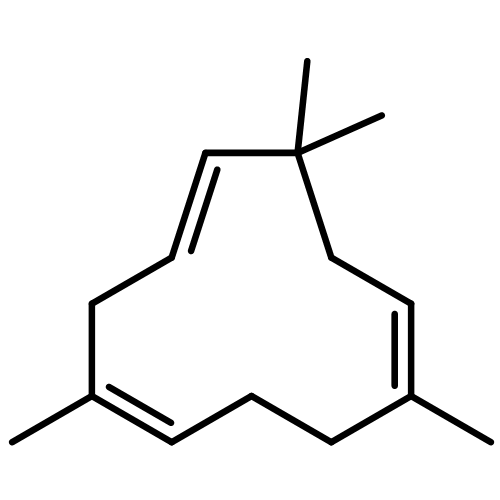



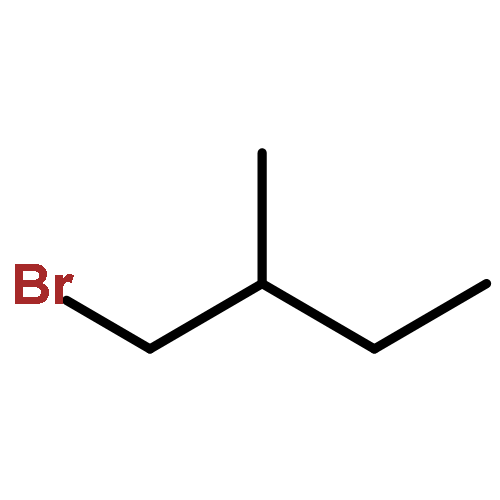


![N-[2-(dimethylamino)ethyl]-4-methylbenzamide](http://img.cochemist.com/ccimg/6000/5925-74-6.png)
![N-[2-(dimethylamino)ethyl]-4-methylbenzamide](http://img.cochemist.com/ccimg/6000/5925-74-6_b.png)
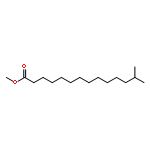





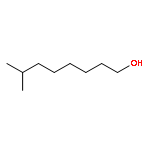









![(1AR,4R,4AR,7S,7AS,7BR)-1,1,4,7-TETRAMETHYLDECAHYDRO-1H-CYCLOPROP<WBR />A[E]AZULENE-4,7-DIOL](http://img.cochemist.com/ccimg/800/763-10-0.png)
![(1AR,4R,4AR,7S,7AS,7BR)-1,1,4,7-TETRAMETHYLDECAHYDRO-1H-CYCLOPROP<WBR />A[E]AZULENE-4,7-DIOL](http://img.cochemist.com/ccimg/800/763-10-0_b.png)







![(3Z)-3-[5-(5-HYDROXY-1H-INDOL-3-YL)-2-OXO-1H-PYRROL-3-YLIDENE]-1H-INDOL-2-ONE](http://img.cochemist.com/ccimg/600/548-54-9.png)
![(3Z)-3-[5-(5-HYDROXY-1H-INDOL-3-YL)-2-OXO-1H-PYRROL-3-YLIDENE]-1H-INDOL-2-ONE](http://img.cochemist.com/ccimg/600/548-54-9_b.png)



![6-METHYL-4-NITRO-1-(PHENYLSULFONYL)-1H-PYRROLO[2,3-B]PYRIDINE](http://img.cochemist.com/ccimg/500/495-60-3.png)
![6-METHYL-4-NITRO-1-(PHENYLSULFONYL)-1H-PYRROLO[2,3-B]PYRIDINE](http://img.cochemist.com/ccimg/500/495-60-3_b.png)
![Tricyclo[6.3.1.02,5]dodecan-1-ol,4,4,8-trimethyl-, (1R,2S,5R,8S)-](http://img.cochemist.com/ccimg/500/472-97-9.png)
![Tricyclo[6.3.1.02,5]dodecan-1-ol,4,4,8-trimethyl-, (1R,2S,5R,8S)-](http://img.cochemist.com/ccimg/500/472-97-9_b.png)
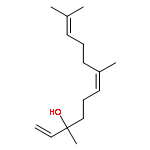
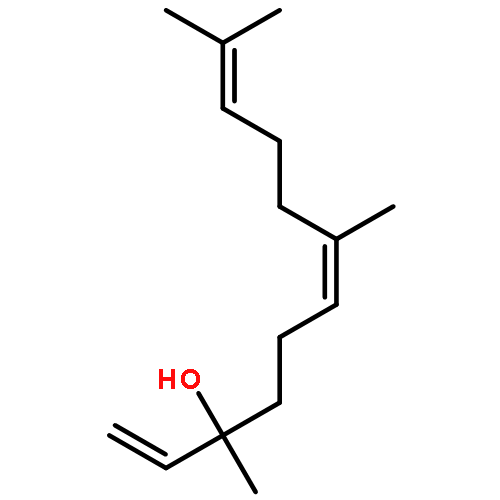


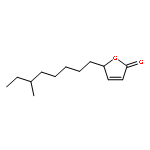













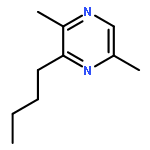
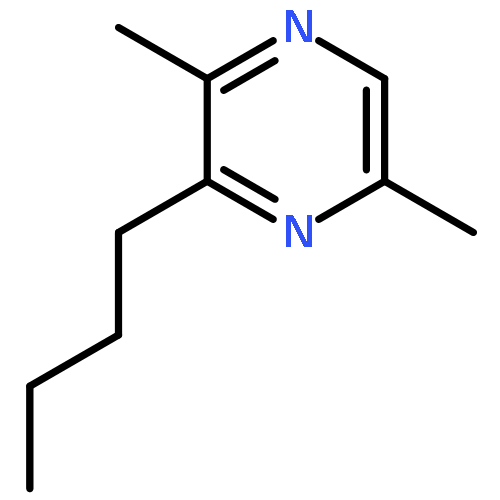




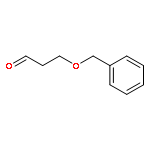
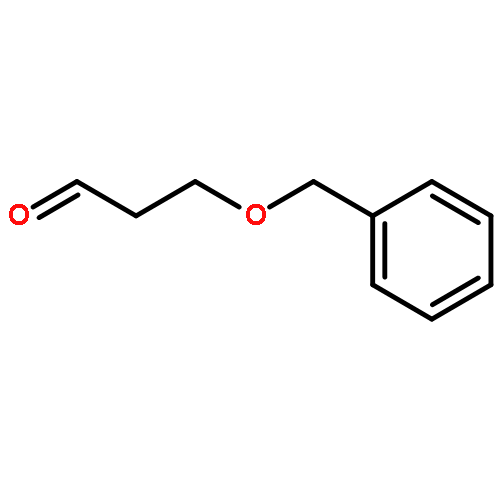


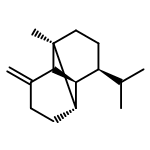







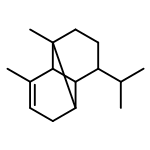







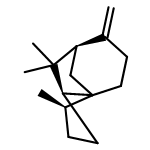
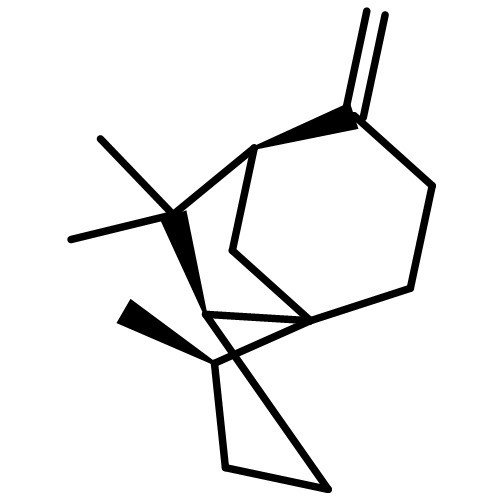


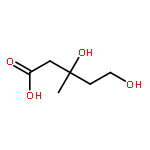

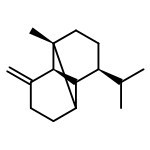
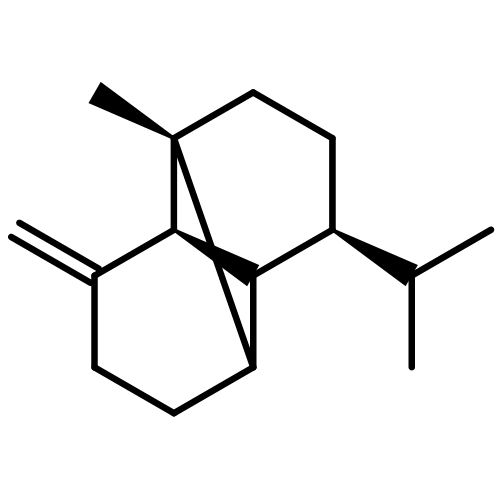
![Cyclohexene, 3-(1,5-dimethyl-4-hexenyl)-6-methylene-, [S-(R*,S*)]-](http://img.cochemist.com/ccimg/20400/20307-83-9.png)
![Cyclohexene, 3-(1,5-dimethyl-4-hexenyl)-6-methylene-, [S-(R*,S*)]-](http://img.cochemist.com/ccimg/20400/20307-83-9_b.png)




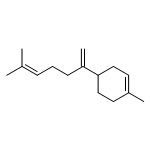
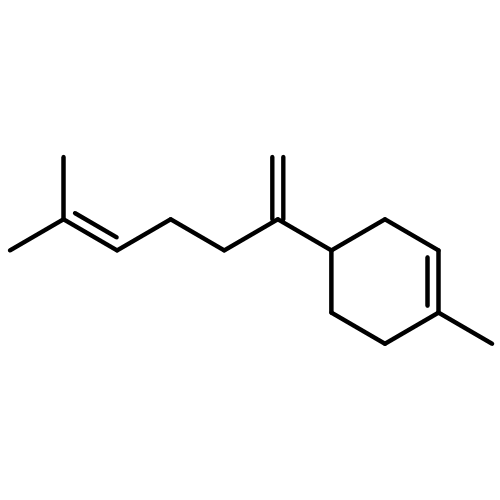


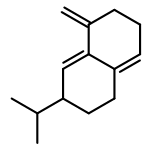
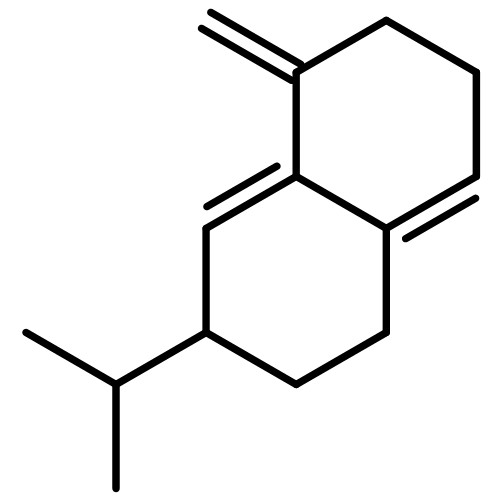
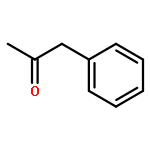
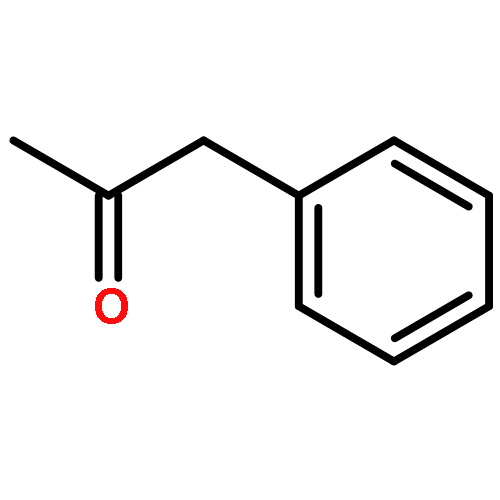



![Bicyclo[2.2.1]heptan-2-ol,1,2,7,7-tetramethyl-, (1R,2R,4R)-rel-](http://img.cochemist.com/ccimg/2400/2371-42-8.png)
![Bicyclo[2.2.1]heptan-2-ol,1,2,7,7-tetramethyl-, (1R,2R,4R)-rel-](http://img.cochemist.com/ccimg/2400/2371-42-8_b.png)