Co-reporter:Carol Ann Castaneda, Jeffrey E. Lopez, Caleb G. Joseph, Michael D. Scholle, Milan Mrksich, and Carol A. Fierke
Biochemistry October 24, 2017 Volume 56(Issue 42) pp:5663-5663
Publication Date(Web):September 22, 2017
DOI:10.1021/acs.biochem.7b00851
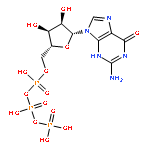
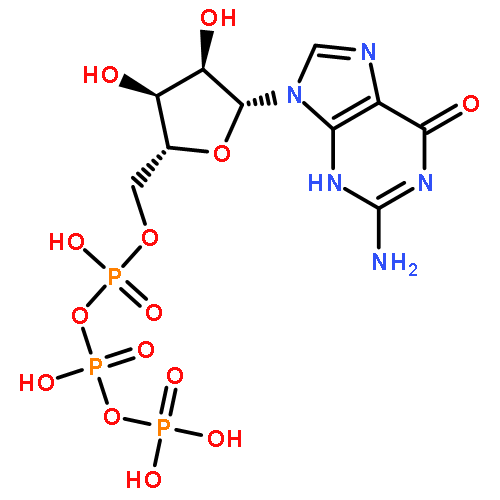
Histone deacetylase 8 (HDAC8) is a well-characterized member of the class I acetyl-lysine deacetylase (HDAC) family. Previous work has shown that the efficiency of HDAC8-catalyzed deacetylation of a methylcoumarin peptide varies depending on the identity of the divalent metal ion in the HDAC8 active site. Here we demonstrate that both HDAC8 activity and substrate selectivity for a diverse range of peptide substrates depend on the identity of the active site metal ion. Varied deacetylase activities of Fe(II)- and Zn(II)-HDAC8 toward an array of peptide substrates were identified using self-assembled monolayers for matrix-assisted laser desorption ionization (SAMDI) mass spectrometry. Subsequently, the metal dependence of deacetylation of peptides of biological interest was measured using an in vitro peptide assay. While Fe(II)-HDAC8 is generally more active than Zn(II)-HDAC8, the Fe(II)/Zn(II) HDAC8 activity ratio varies widely (from 2 to 150) among the peptides tested. These data provide support for the hypothesis that HDAC8 may undergo metal switching in vivo that, in turn, may regulate its activity. However, future studies are needed to explore the identity of the metal ion bound to HDAC8 in cells under varied conditions.
Co-reporter:Jeffrey E. Lopez, Sarah E. Haynes, Jaimeen D. Majmudar, Brent R. Martin, and Carol A. Fierke
Journal of the American Chemical Society November 15, 2017 Volume 139(Issue 45) pp:16222-16222
Publication Date(Web):October 16, 2017
DOI:10.1021/jacs.7b07603
The histone deacetylase family comprises 18 enzymes that catalyze deacetylation of acetylated lysine residues; however, the specificity and substrate profile of each isozyme remains largely unknown. Due to transient enzyme–substrate interactions, conventional co-immunoprecipitation methods frequently fail to identify enzyme-specific substrates. Additionally, compensatory mechanisms often limit the ability of knockdown or chemical inhibition studies to achieve significant fold changes observed by acetylation proteomics methods. Furthermore, measured alterations do not guarantee a direct link between enzyme and substrate. Here we present a chemical crosslinking strategy that incorporates a photoreactive, non-natural amino acid, p-benzoyl-l-phenylalanine, into various positions of the structurally characterized isozyme histone deacetylase 8 (HDAC8). After covalent capture, co-immunoprecipitation, and mass spectrometric analysis, we identified a subset of HDAC8 substrates from human cell lysates, which were further validated for catalytic turnover. Overall, this chemical crosslinking approach identified novel HDAC8-specific substrates with high catalytic efficiency, thus presenting a general strategy for unbiased deacetylase substrate discovery.
Co-reporter:Jeffrey E. López, Eric D. Sullivan, and Carol A. Fierke
ACS Chemical Biology 2016 Volume 11(Issue 3) pp:706
Publication Date(Web):February 24, 2016
DOI:10.1021/acschembio.5b01067
Epigenetic regulation is a key factor in cellular homeostasis. Post-translational modifications (PTMs) are a central focus of this regulation as they function as signaling markers within the cell. Lysine acetylation is a dynamic, reversible PTM that has garnered recent attention due to alterations in various types of cancer. Acetylation levels are regulated by two opposing enzyme families: lysine acetyltransferases (KATs) and histone deacetylases (HDACs). HDACs are key players in epigenetic regulation and have a role in the silencing of tumor suppressor genes. The dynamic equilibrium of acetylation makes HDACs attractive targets for drug therapy. However, substrate selectivity and biological function of HDAC isozymes is poorly understood. This review outlines the current understanding of the roles and specific epigenetic interactions of the metal-dependent HDACs in addition to their roles in cancer.
Co-reporter:Sister M. Lucy Gantt FSGM, Christophe Decroos, Matthew S. Lee, Laura E. Gullett, Christine M. Bowman, David W. Christianson, and Carol A. Fierke
Biochemistry 2016 Volume 55(Issue 5) pp:820-832
Publication Date(Web):January 25, 2016
DOI:10.1021/acs.biochem.5b01327
Histone deacetylases (HDACs) regulate cellular processes such as differentiation and apoptosis and are targeted by anticancer therapeutics in development and in the clinic. HDAC8 is a metal-dependent class I HDAC and is proposed to use a general acid–base catalytic pair in the mechanism of amide bond hydrolysis. Here, we report site-directed mutagenesis and enzymological measurements to elucidate the catalytic mechanism of HDAC8. Specifically, we focus on the catalytic function of Y306 and the histidine-aspartate dyads H142-D176 and H143-D183. Additionally, we report X-ray crystal structures of four representative HDAC8 mutants: D176N, D176N/Y306F, D176A/Y306F, and H142A/Y306F. These structures provide a useful framework for understanding enzymological measurements. The pH dependence of kcat/KM for wild-type Co(II)-HDAC8 is bell-shaped with two pKa values of 7.4 and 10.0. The upper pKa reflects the ionization of the metal-bound water molecule and shifts to 9.1 in Zn(II)-HDAC8. The H142A mutant has activity 230-fold lower than that of wild-type HDAC8, but the pKa1 value is not altered. Y306F HDAC8 is 150-fold less active than the wild-type enzyme; crystal structures show that Y306 hydrogen bonds with the zinc-bound substrate carbonyl, poised for transition state stabilization. The H143A and H142A/H143A mutants exhibit activity that is >80000-fold lower than that of wild-type HDAC8; the buried D176N and D176A mutants have significant catalytic effects, with more subtle effects caused by D183N and D183A. These enzymological and structural studies strongly suggest that H143 functions as a single general base–general acid catalyst, while H142 remains positively charged and serves as an electrostatic catalyst for transition state stabilization.
Co-reporter:Benjamin C. Jennings, Amy M. Danowitz, Yen-Chih Wang, Richard A. Gibbs, Mark D. Distefano, Carol A. Fierke
Bioorganic & Medicinal Chemistry Letters 2016 Volume 26(Issue 4) pp:1333-1336
Publication Date(Web):15 February 2016
DOI:10.1016/j.bmcl.2015.12.079
Attempts to identify the prenyl-proteome of cells or changes in prenylation following drug treatment have used ‘clickable’ alkyne-modified analogs of the lipid substrates farnesyl- and geranylgeranyl-diphosphate (FPP and GGPP). We characterized the reactivity of four alkyne-containing analogs of FPP with purified protein farnesyltransferase and a small library of dansylated peptides using an in vitro continuous spectrofluorimetric assay. These analogs alter prenylation specificity and reactivity suggesting that in vivo results obtained using these FPP analogs should be interpreted cautiously.
Co-reporter:Kevin B. Daniel; Eric D. Sullivan; Yao Chen; Joshua C. Chan; Patricia A. Jennings; Carol A. Fierke;Seth M. Cohen
Journal of Medicinal Chemistry 2015 Volume 58(Issue 11) pp:4812-4821
Publication Date(Web):May 14, 2015
DOI:10.1021/acs.jmedchem.5b00539
Histone deacetylase inhibitors (HDACi) target abnormal epigenetic states associated with a variety of pathologies, including cancer. Here, the development of a prodrug of the canonical broad-spectrum HDACi suberoylanilide hydroxamic acid (SAHA) is described. Although hydroxamic acids are utilized universally in the development of metalloenzyme inhibitors, they are considered to be poor pharmacophores with reduced activity in vivo. We developed a prodrug of SAHA by appending a promoiety, sensitive to thiols, to the hydroxamic acid warhead (termed SAHA-TAP). After incubation of SAHA-TAP with an HDAC, the thiol of a conserved HDAC cysteine residue becomes covalently tagged with the promoiety, initiating a cascade reaction that leads to the release of SAHA. Mass spectrometry and enzyme kinetics experiments validate that the cysteine residue is covalently appended with the TAP promoiety. SAHA-TAP demonstrates cytotoxicity activity against various cancer cell lines. This strategy represents an original prodrug design with a dual mode of action for HDAC inhibition.
Co-reporter:Noah A. Wolfson, Carol Ann Pitcairn, Eric D. Sullivan, Caleb G. Joseph, Carol A. Fierke
Analytical Biochemistry 2014 Volume 456() pp:61-69
Publication Date(Web):1 July 2014
DOI:10.1016/j.ab.2014.03.012
Abstract
Histone deacetylases catalyze the hydrolysis of an acetyl group from post-translationally modified acetyl-lysine residues in a wide variety of essential cellular proteins, including histones. Because these lysine modifications can alter the activity and properties of affected proteins, aberrant acetylation/deacetylation may contribute to disease states. Many fundamental questions regarding the substrate specificity and regulation of these enzymes have yet to be answered. Here, we optimize an enzyme-coupled assay to measure low micromolar concentrations of acetate, coupling acetate production to the formation of NADH (nicotinamide adenine dinucleotide, reduced form) that is measured by changes in either absorbance or fluorescence. Using this assay, we measured the steady-state kinetics of peptides representing the H4 histone tail and demonstrate that a C-terminally conjugated methylcoumarin enhances the catalytic efficiency of deacetylation catalyzed by cobalt(II)-bound histone deacetylase 8 [Co(II)–HDAC8] compared with peptide substrates containing a C-terminal carboxylate, amide, and tryptophan by 50-, 2.8-, and 2.3-fold, respectively. This assay can be adapted for a high-throughput screening format to identify HDAC substrates and inhibitors.
Co-reporter:Ningkun Wang;Jean M. Lodge;Anna K. Mapp
PNAS 2014 Volume 111 (Issue 33 ) pp:12061-12066
Publication Date(Web):2014-08-19
DOI:10.1073/pnas.1406033111
Allosteric binding events play a critical role in the formation and stability of transcriptional activator–coactivator complexes,
perhaps in part due to the often intrinsically disordered nature of one or more of the constituent partners. The kinase-inducible
domain interacting (KIX) domain of the master coactivator CREB binding protein/p300 is a conformationally dynamic domain that
complexes with transcriptional activators at two discrete binding sites in allosteric communication. The complexation of KIX
with the transcriptional activation domain of mixed-lineage leukemia protein leads to an enhancement of binding by the activation
domain of CREB (phosphorylated kinase-inducible domain of CREB) to the second site. A transient kinetic analysis of the ternary
complex formation aided by small molecule ligands that induce positive or negative cooperative binding reveals that positive
cooperativity is largely governed by stabilization of the bound complex as indicated by a decrease in koff. Thus, this suggests the increased binding affinity for the second ligand is not due to an allosteric creation of a more
favorable binding interface by the first ligand. This is consistent with data from us and from others indicating that the
on rates of conformationally dynamic proteins approach the limits of diffusion. In contrast, negative cooperativity is manifested
by alterations in both kon and koff, suggesting stabilization of the binary complex.
Co-reporter:Da Wang and Carol A. Fierke
Metallomics 2013 vol. 5(Issue 4) pp:372-383
Publication Date(Web):27 Feb 2013
DOI:10.1039/C3MT20217H
Intracellular zinc homeostasis is regulated by an extensive network of transporters, ligands and transcription factors. The zinc detoxification functions of three transporters and a periplasmic protein regulated by the BaeSR two-component system were explored in this work by evaluating the effect of single gene knockouts in the BaeSR regulon on the cell growth rate, free zinc, total zinc and total copper after zinc shock. Two exporters, MdtABC and MdtD, and the periplasmic protein, Spy, are involved in zinc detoxification based on the growth defects at high cell density and increases in free (>1000-fold) and total zinc/copper (>2-fold) that were observed in the single knockout strains upon exposure to zinc. These proteins complement the ATP-driven zinc export mediated by ZntA in E. coli to limit zinc toxicity. These results highlight the functions of the BaeSR regulon in metal homeostasis.
Co-reporter:Noah A. Wolfson;Carol Ann Pitcairn
Biopolymers 2013 Volume 99( Issue 2) pp:112-126
Publication Date(Web):
DOI:10.1002/bip.22135
Abstract
The lysine deacetylase family of enzymes (HDACs) was first demonstrated to catalyze deacetylation of acetyllysine residues on histones. In subsequent years, HDACs have been shown to recognize a large pool of acetylated nonhistone proteins as substrates. Recently, thousands of acetylated proteins have been discovered, yet in most cases, the HDAC that catalyzes deacetylation in vivo has not been identified. This gap has created the need for better in vivo, in vitro, and in silico approaches for determining HDAC substrates. While HDAC8 is the best kinetically and structurally characterized HDAC, few efficient substrates have yet been substantiated in vivo. In this review, we delineate factors that may be important for determining HDAC8 substrate recognition and catalytic activity, including structure, complex formation, and post-translational modifications. This summary provides insight into the challenges of identifying in vivo substrates for HDAC8, and provides a good vantage point for understanding the variables important for predicting HDAC substrate recognition. © 2012 Wiley Periodicals, Inc. Biopolymers 99: 112–126, 2013.
Co-reporter:Elaina A Zverina, Corissa L Lamphear, Elia N Wright, Carol A Fierke
Current Opinion in Chemical Biology 2012 Volume 16(5–6) pp:544-552
Publication Date(Web):December 2012
DOI:10.1016/j.cbpa.2012.10.015
Protein post-translational modifications increase the functional diversity of the proteome by covalently adding chemical moieties onto proteins thereby changing their activation state, cellular localization, interacting partners, and life cycle. Lipidation is one such modification that enables membrane association of naturally cytosolic proteins. Protein prenyltransferases irreversibly install isoprenoid units of varying length via a thioether linkage onto proteins that exert their cellular activity at membranes. Substrates of prenyltransferases are involved in countless signaling pathways and processes within the cell. Identification of new prenylation substrates, prenylation pathway regulators, and dynamic trafficking of prenylated proteins are all avenues of intense, ongoing research that are challenging, exciting, and have the potential to significantly advance the field in the near future.Graphical abstractHighlights► Expansion of the CaaX paradigm by biochemical and computational methodologies. ► Improvement of chemical biology tools to define mammalian prenylome. ► Identification of proteins that regulate entry into prenylation pathway. ► Involvement of mammalian prenyltransferases in bacterial pathogenesis. ► Clinical evaluation of inhibitors for cancer and premature aging diseases.
Co-reporter:Manoj Cheriyan, Eric J. Toone, and Carol A. Fierke
Biochemistry 2012 Volume 51(Issue 8) pp:
Publication Date(Web):February 1, 2012
DOI:10.1021/bi201899b
The substrate specificity of enzymes is frequently narrow and constrained by multiple interactions, limiting the use of natural enzymes in biocatalytic applications. Aldolases have important synthetic applications, but the usefulness of these enzymes is hampered by their narrow reactivity profile with unnatural substrates. To explore the determinants of substrate selectivity and alter the specificity of Escherichia coli 2-keto-3-deoxy-6-phosphogluconate (KDPG) aldolase, we employed structure-based mutagenesis coupled with library screening of mutant enzymes localized to the bacterial periplasm. We identified two active site mutations (T161S and S184L) that work additively to enhance the substrate specificity of this aldolase to include catalysis of retro-aldol cleavage of (4S)-2-keto-4-hydroxy-4-(2′-pyridyl)butyrate (S-KHPB). These mutations improve the value of kcat/KMS-KHPB by >450-fold, resulting in a catalytic efficiency that is comparable to that of the wild-type enzyme with the natural substrate while retaining high stereoselectivity. Moreover, the value of kcatS-KHPB for this mutant enzyme, a parameter critical for biocatalytic applications, is 3-fold higher than the maximal value achieved by the natural aldolase with any substrate. This mutant also possesses high catalytic efficiency for the retro-aldol cleavage of the natural substrate, KDPG, and a >50-fold improved activity for cleavage of 2-keto-4-hydroxy-octonoate, a nonfunctionalized hydrophobic analogue. These data suggest a substrate binding mode that illuminates the origin of facial selectivity in aldol addition reactions catalyzed by KDPG and 2-keto-3-deoxy-6-phosphogalactonate aldolases. Furthermore, targeting mutations to the active site provides a marked improvement in substrate selectivity, demonstrating that structure-guided active site mutagenesis combined with selection techniques can efficiently identify proteins with characteristics that compare favorably to those of naturally occurring enzymes.
Co-reporter:Da Wang, Olijahwon Hosteen, Carol A. Fierke
Journal of Inorganic Biochemistry 2012 Volume 111() pp:173-181
Publication Date(Web):June 2012
DOI:10.1016/j.jinorgbio.2012.02.008
In E. coli, ZitB and ZntA are important metal exporters that enhance cell viability under high environmental zinc. To understand their functions in maintaining zinc homeostasis, we applied a novel genetically-encoded fluorescent zinc sensor to monitor the intracellular free zinc changes in wild type, ∆zitB and ∆zntA E. coli cells upon sudden exposure to toxic levels of zinc (“zinc shock”). The intracellular readily exchangeable zinc concentration (or “free” zinc) increases transiently from picomolar to nanomolar levels, accelerating zinc-activated gene transcription. After zinc shock, the zitB mRNA level is constant while the zntA mRNA increases substantially in a zinc-dependent manner. In the ∆zitB E. coli strain the free zinc concentration rises more rapidly after zinc shock compared to wild type cells while a prolonged accumulation of free zinc is observed in the ∆zntA strain. Based on these results, we propose that ZitB functions as a constitutive, first-line defense against toxic zinc influx, while ZntA is up-regulated to efficiently lower the free zinc concentration. Furthermore, the ZntR-mediated transcription of zntA exhibits an apparent K1/2 for zinc activation in the nanomolar range in vivo, significantly higher than the femtomolar affinity for zinc binding and transcription activation previously measured in vitro. A kinetically-controlled transcription model is sufficient to explain the observed regulation of intracellular free zinc concentration by ZntR and ZntA after zinc shock.Intracellular free zinc rises to nanomolar range in E. coli upon zinc shock, activating the ZntR-mediated zntA transcription.Highlights► Sudden exposure to high zinc causes transient increases in free zinc in E. coli. ► Intracellular free zinc peak accelerates zinc-activated transcription. ► ZntR-mediated zntA transcription has nanomolar K1/2 of zinc. ► ZitB acts as the first line defense against high zinc. ►Detoxification of excess zinc requires the expression of zntA.
Co-reporter:Wan Hsin Lim;Michael J. Howard;Markos Koutmos
PNAS 2012 Volume 109 (Issue 40 ) pp:16149-16154
Publication Date(Web):2012-10-02
DOI:10.1073/pnas.1209062109
Ribonuclease P (RNase P) catalyzes the maturation of the 5′ end of tRNA precursors. Typically these enzymes are ribonucleoproteins
with a conserved RNA component responsible for catalysis. However, protein-only RNase P (PRORP) enzymes process precursor
tRNAs in human mitochondria and in all tRNA-using compartments of Arabidopsis thaliana. PRORP enzymes are nuclear encoded and conserved among many eukaryotes, having evolved recently as yeast mitochondrial genomes
encode an RNase P RNA. Here we report the crystal structure of PRORP1 from A. thaliana at 1.75 Å resolution, revealing a prototypical metallonuclease domain tethered to a pentatricopeptide repeat (PPR) domain
by a structural zinc-binding domain. The metallonuclease domain is a unique high-resolution structure of a Nedd4-BP1, YacP
Nucleases (NYN) domain that is a member of the PIN domain-like fold superfamily, including the FLAP nuclease family. The structural
similarity between PRORP1 and the FLAP nuclease family suggests that they evolved from a common ancestor. Biochemical data
reveal that conserved aspartate residues in PRORP1 are important for catalytic activity and metal binding and that the PPR
domain also enhances activity, likely through an interaction with pre-tRNA. These results provide a foundation for understanding
tRNA maturation in organelles. Furthermore, these studies allow for a molecular-level comparison of the catalytic strategies
used by the only known naturally evolved protein and RNA-based catalysts that perform the same biological function, pre-tRNA
maturation, thereby providing insight into the differences between the prebiotic RNA world and the present protein-dominated
world.
Co-reporter:Manoj Cheriyan, Matthew J. Walters, Brian D. Kang, Laura L. Anzaldi, Eric J. Toone, Carol A. Fierke
Bioorganic & Medicinal Chemistry 2011 Volume 19(Issue 21) pp:6447-6453
Publication Date(Web):1 November 2011
DOI:10.1016/j.bmc.2011.08.056
The use of biological catalysts for industrial scale synthetic chemistry is highly attractive, given their cost effectiveness, high specificity that obviates the need for protecting group chemistry, and the environmentally benign nature of enzymatic procedures. Here we evolve the naturally occurring 2-keto-3-deoxy-6-phosphogluconate (KDPG) aldolases from Thermatoga maritima and Escherichia coli, into enzymes that recognize a nonfunctionalized electrophilic substrate, 2-keto-4-hydroxyoctonoate (KHO). Using an in vivo selection based on pyruvate auxotrophy, mutations were identified that lower the KM value up to 100-fold in E. coli KDPG aldolase, and that enhance the efficiency of retro-aldol cleavage of KHO by increasing the value of kcat/KM up to 25-fold in T. maritima KDPG aldolase. These data indicate that numerous mutations distal from the active site contribute to enhanced ‘uniform binding’ of the substrates, which is the first step in the evolution of novel catalytic activity.
Co-reporter:XiaoMu Guan
Science China Chemistry 2011 Volume 54( Issue 12) pp:1888-1897
Publication Date(Web):2011 December
DOI:10.1007/s11426-011-4428-2
Protein palmitoylation is a widespread lipid modification in which one or more cysteine thiols on a substrate protein are modified to form a thioester with a palmitoyl group. This lipid modification is readily reversible; a feature of protein palmitoylation that allows for rapid regulation of the function of many cellular proteins. Mutations in palmitoyltransferases (PATs), the enzymes that catalyze the formation of this modification, are associated with a number of neurological diseases and cancer progression. This review summarizes the crucial role of palmitoylation in biological systems, the discovery of the DHHC protein family that catalyzes protein palmitoylation, and the development of methods for investigating the catalytic mechanism of PATs.
Co-reporter:Marcy Hernick, Samuel G. Gattis, James E. Penner-Hahn and Carol A. Fierke
Biochemistry 2010 Volume 49(Issue 10) pp:
Publication Date(Web):February 5, 2010
DOI:10.1021/bi902066t
The metal-dependent deacetylase UDP-3-O-[(R)-3-hydroxymyristoyl]-N-acetylglucosamine deacetylase (LpxC) catalyzes the first committed step in lipid A biosynthesis, the hydrolysis of UDP-3-O-myristoyl-N-acetylglucosamine to form UDP-3-O-myristoylglucosamine and acetate. Consequently, LpxC is a target for the development of antibiotics, nearly all of which coordinate the active site metal ion. Here we examine the ability of Fe2+ to serve as a cofactor for wild-type Escherichia coli LpxC and a mutant enzyme (EcC63A), in which one of the ligands for the inhibitory metal binding site has been removed. LpxC exhibits higher activity (6−8-fold) with a single bound Fe2+ as the cofactor compared to Zn2+-LpxC; both metalloenzymes have a bell-shaped dependence on pH with similar pKa values, indicating that at least two ionizations are important for maximal activity. X-ray absorption spectroscopy experiments suggest that the catalytic metal ion bound to Fe2+-EcLpxC is five-coordinate, suggesting that catalytic activity may correlate with coordination number. Furthermore, the ligand affinity of Fe2+-LpxC compared to the Zn2+ enzyme is altered by up to 6-fold. In contrast to Zn2+-LpxC, the activity of Fe2+-LpxC is redox-sensitive, and a time-dependent decrease in activity is observed under aerobic conditions. The LpxC activity of crude E. coli cell lysates is also aerobically sensitive, consistent with the presence of Fe2+-LpxC. These data indicate that EcLpxC can use either Fe2+ or Zn2+ to activate catalysis in vitro and possibly in vivo, which may allow LpxC to function in E. coli grown under different environmental conditions.
Co-reporter:Tamiika K. Hurst, Da Wang, Richard B. Thompson, Carol A. Fierke
Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics 2010 Volume 1804(Issue 2) pp:393-403
Publication Date(Web):February 2010
DOI:10.1016/j.bbapap.2009.09.031
Carbonic anhydrases are archetypical zinc metalloenzymes and as such, they have been developed as the recognition element of a family of fluorescent indicators (sensors) to detect metal ions, particularly Zn2+ and Cu2+. Subtle modification of the structure of human carbonic anhydrase II isozyme (CAII) alters the selectivity, sensitivity, and response time for these sensors. Sensors using CAII variants coupled with zinc-dependent fluorescent ligands demonstrate picomolar sensitivity, unmatched selectivity, ratiometric fluorescence signal, and near diffusion-controlled response times. Recently, these sensors have been applied to measuring the readily exchangeable concentrations of zinc in the cytosol and nucleus of mammalian tissue culture cells and concentrations of free Cu2+ in seawater.
Co-reporter:Kristin S. Koutmou;Anette Casiano-Negroni;James E. Penner-Hahn;Hashim M. Al-Hashimi;Samuel Pazicni;Melissa M. Getz;Andrew J. Andrews
PNAS 2010 Volume 107 (Issue 6 ) pp:2479-2484
Publication Date(Web):2010-02-09
DOI:10.1073/pnas.0906319107
Functionally critical metals interact with RNA through complex coordination schemes that are currently difficult to visualize
at the atomic level under solution conditions. Here, we report a new approach that combines NMR and XAS to resolve and characterize
metal binding in the most highly conserved P4 helix of ribonuclease P (RNase P), the ribonucleoprotein that catalyzes the
divalent metal ion-dependent maturation of the 5′ end of precursor tRNA. Extended X-ray absorption fine structure (EXAFS)
spectroscopy reveals that the Zn2+ bound to a P4 helix mimic is six-coordinate, with an average Zn-O/N bond distance of 2.08 Å. The EXAFS data also show intense
outer-shell scattering indicating that the zinc ion has inner-shell interactions with one or more RNA ligands. NMR Mn2+ paramagnetic line broadening experiments reveal strong metal localization at residues corresponding to G378 and G379 in B. subtilis RNase P. A new “metal cocktail” chemical shift perturbation strategy involving titrations with
, Zn2+, and
confirm an inner-sphere metal interaction with residues G378 and G379. These studies present a unique picture of how metals
coordinate to the putative RNase P active site in solution, and shed light on the environment of an essential metal ion in
RNase P. Our experimental approach presents a general method for identifying and characterizing inner-sphere metal ion binding
sites in RNA in solution.
Co-reporter:James L. Hougland, Corissa L. Lamphear, Sarah A. Scott, Richard A. Gibbs and Carol A. Fierke
Biochemistry 2009 Volume 48(Issue 8) pp:
Publication Date(Web):February 6, 2009
DOI:10.1021/bi801710g
Prenylation is a posttranslational modification whereby C-terminal lipidation leads to protein localization to membranes. A C-terminal “Ca1a2X” sequence has been proposed as the recognition motif for two prenylation enzymes, protein farnesyltransferase (FTase) and protein geranylgeranyltransferase type I. To define the parameters involved in recognition of the a2 residue, we performed structure−activity analysis which indicates that FTase discriminates between peptide substrates based on both the hydrophobicity and steric volume of the side chain at the a2 position. For nonpolar side chains, the dependence of the reactivity on side chain volume at this position forms a pyramidal pattern with a maximal activity near the steric volume of valine. This discrimination occurs at a step in the kinetic mechanism that is at or before the farnesylation step. Furthermore, a2 selectivity is also affected by the identity of the adjacent X residue, leading to context-dependent substrate recognition. Context-dependent a2 selectivity suggests that FTase recognizes the sequence downstream of the conserved cysteine as a set of two or three cooperative, interconnected recognition elements as opposed to three independent amino acids. These findings expand the pool of proposed FTase substrates in cells. A better understanding of the molecular recognition of substrates performed by FTase will aid in both designing new FTase inhibitors as therapeutic agents and characterizing proteins involved in prenylation-dependent cellular pathways.
Co-reporter:Matthew J. Walters, Velupillai Srikannathasan, Andrew R. McEwan, James H. Naismith, Carol A. Fierke, Eric J. Toone
Bioorganic & Medicinal Chemistry 2008 Volume 16(Issue 2) pp:710-720
Publication Date(Web):15 January 2008
DOI:10.1016/j.bmc.2007.10.043
2-Keto-3-deoxy-6-phosphogluconate (KDPG) and 2-keto-3-deoxy-6-phosphogalactonate (KDPGal) aldolases catalyze an identical reaction differing in substrate specificity in only the configuration of a single stereocenter. However, the proteins show little sequence homology at the amino acid level. Here we investigate the determinants of substrate selectivity of these enzymes. The Escherichia coli KDPGal aldolase gene, cloned into a T7 expression vector and overexpressed in E. coli, catalyzes retro-aldol cleavage of the natural substrate, KDPGal, with values of kcat/KM and kcat of 1.9 × 104 M−1 s−1 and 4 s−1, respectively. In the synthetic direction, KDPGal aldolase efficiently catalyzes an aldol addition using a limited number of aldehyde substrates, including d-glyceraldehyde-3-phosphate (natural substrate), d-glyceraldehyde, glycolaldehyde, and 2-pyridinecarboxaldehyde. A preparative scale reaction between 2-pyridinecarboxaldehyde and pyruvate catalyzed by KDPGal aldolase produced the aldol adduct of the R stereochemistry in >99.7% ee, a result complementary to that observed using the related KDPG aldolase. The native crystal structure has been solved to a resolution of 2.4 Å and displays the same (α/β)8 topology, as KDPG aldolase. We have also determined a 2.1 Å structure of a Schiff base complex between the enzyme and its substrate. This model predicts that a single amino acid change, T161 in KDPG aldolase to V154 in KDPGal aldolase, plays an important role in determining the stereochemical course of enzyme catalysis and this prediction was borne out by site-directed mutagenesis studies. However, additional changes in the enzyme sequence are required to prepare an enzyme with both high catalytic efficiency and altered stereochemistry.The crystal structure of the E. coli KDPGal aldolase, which displays (α/β)8 topology, has been solved. In addition, site-directed mutagenesis has been employed to investigate the stereoselectivity compared to the analogous KDPG aldolases.
Co-reporter:Stephen W.B. Fullerton, Jennifer S. Griffiths, Alexandra B. Merkel, Manoj Cheriyan, Nathan J. Wymer, Michael J. Hutchins, Carol A. Fierke, Eric J. Toone, James H. Naismith
Bioorganic & Medicinal Chemistry 2006 Volume 14(Issue 9) pp:3002-3010
Publication Date(Web):1 May 2006
DOI:10.1016/j.bmc.2005.12.022
In vivo, 2-keto-3-deoxy-6-phosphogluconate (KDPG) aldolase catalyzes the reversible, stereospecific retro-aldol cleavage of KDPG to pyruvate and d-glyceraldehyde-3-phosphate. The enzyme is a lysine-dependent (Class I) aldolase that functions through the intermediacy of a Schiff base. Here, we propose a mechanism for this enzyme based on crystallographic studies of wild-type and mutant aldolases. The three dimensional structure of KDPG aldolase from the thermophile Thermotoga maritima was determined to 1.9 Å. The structure is the standard α/β barrel observed for all Class I aldolases. At the active site Lys we observe clear density for a pyruvate Schiff base. Density for a sulfate ion bound in a conserved cluster of residues close to the Schiff base is also observed. We have also determined the structure of a mutant of Escherichia coli KDPG aldolase in which the proposed general acid/base catalyst has been removed (E45N). One subunit of the trimer contains density suggesting a trapped pyruvate carbinolamine intermediate. All three subunits contain a phosphate ion bound in a location effectively identical to that of the sulfate ion bound in the T. maritima enzyme. The sulfate and phosphate ions experimentally locate the putative phosphate binding site of the aldolase and, together with the position of the bound pyruvate, facilitate construction of a model for the full-length KDPG substrate complex. The model requires only minimal positional adjustments of the experimentally determined covalent intermediate and bound anion to accommodate full-length substrate. The model identifies the key catalytic residues of the protein and suggests important roles for two observable water molecules. The first water molecule remains bound to the enzyme during the entire catalytic cycle, shuttling protons between the catalytic glutamate and the substrate. The second water molecule arises from dehydration of the carbinolamine and serves as the nucleophilic water during hydrolysis of the enzyme-product Schiff base. The second water molecule may also mediate the base-catalyzed enolization required to form the carbon nucleophile, again bridging to the catalytic glutamate. Many aspects of this mechanism are observed in other Class I aldolases and suggest a mechanistically and, perhaps, evolutionarily related family of aldolases distinct from the N-acetylneuraminate lyase (NAL) family.A mechanism for catalysis by the Class I aldolase 2-keto-3-deoxy-6-phosphogluconate aldolase is reported based on crystallographic studies of native and mutant variants of both the Thermotoga maritima and Escherichia coli KDPG aldolase.
Co-reporter:James P. Sumner, Nissa M. Westerberg, Andrea K. Stoddard, Tamiika K. Hurst, Michele Cramer, Richard B. Thompson, Carol A. Fierke, Raoul Kopelman
Biosensors and Bioelectronics 2006 Volume 21(Issue 7) pp:1302-1308
Publication Date(Web):15 January 2006
DOI:10.1016/j.bios.2005.04.023
The wild type form of Red fluorescent protein (DsRed), an intrinsically fluorescent protein found in tropical corals, is found to be highly selective, reversible and sensitive for both Cu+ and Cu2+, with a nanomolar detection limit. The selectivity towards these ions is retained even in the presence of other heavy metal ions. The Kd values for monovalent and divalent copper, based on single binding isotherms, are 450 and 540 nM, respectively. The wild type DsRed sensitivity to Cu2+ (below 1 ppb) is seven orders of magnitude better than that of the related wild type Green Fluorescent protein (GFP), and it is even 40 times more sensitive than engineered mutants of GFP. Potential binding sites have been proposed, based on amino acid sequences for copper binding and the distance from the chromophore, with the aid of computer modeling.
Co-reporter:June E. Pais, Katherine E. Bowers, Andrea K. Stoddard, Carol A. Fierke
Analytical Biochemistry 2005 Volume 345(Issue 2) pp:302-311
Publication Date(Web):15 October 2005
DOI:10.1016/j.ab.2005.07.040
Protein farnesyltransferase and protein geranylgeranyltransferase type I catalyze the transfer of a 15- and a 20-carbon prenyl group, respectively, from a prenyl diphosphate to a cysteine residue at the carboxyl terminus of target proteins, with the concomitant release of diphosphate. Common substrates include oncogenic Ras proteins, which are implicated in up to 30% of all human cancers, making prenyltransferases a viable target for chemotherapeutic drugs. A coupled assay has been developed to measure the rate constant of diphosphate (PPi) dissociation during the prenyltransferase reaction under both single and multiple turnover conditions. In this assay, the PPi group produced in the prenyltransferase reaction is rapidly cleaved by inorganic pyrophosphatase to form phosphate (Pi), which is then bound by a coumarin-labeled phosphate binding protein from Escherichia coli, resulting in a fluorescence increase. The observed rate constant for PPi release is equal to the rate constant of prenylation of the peptide, as measured by other assays, so that this nonradioactive assay can be used to measure prenyltransferase activity under either single or multiple turnover conditions. This assay can be adapted for high-throughput screening for potential prenyltransferase substrates and inhibitors.
Co-reporter:Jennifer S Griffiths, Manoj Cheriyan, Jayme B Corbell, Luka Pocivavsek, Carol A Fierke, Eric J Toone
Bioorganic & Medicinal Chemistry 2004 Volume 12(Issue 15) pp:4067-4074
Publication Date(Web):1 August 2004
DOI:10.1016/j.bmc.2004.05.034
A novel bacterial in vivo selection for pyruvate aldolase activity is described. Pyruvate kinase deficient cells, which lack the ability to biosynthetically generate pyruvate, require supplementation of exogenous pyruvate when grown on ribose. Supplementation with pyruvate concentrations as low as 50 μM rescues cell growth. A known substrate of the KDPG aldolases, 2-keto-4-hydroxy-4-(2′-pyridyl)butyrate (KHPB), also rescues cell growth, consistent with retroaldol cleavage by KDPG aldolase and rescue through pyruvate release. An initial round of selection against 2-keto-4-hydroxyoctonate (KHO), a nonsubstrate for wild-type aldolase, produced three mutants with intriguing alterations in protein sequence. This selection system allows rapid screening of mutant enzyme libraries and facilitates the discovery of enzymes with novel substrate specificities.Graphic
Co-reporter:John Hsieh;Andy J. Andrews
Biopolymers 2004 Volume 73(Issue 1) pp:
Publication Date(Web):8 DEC 2003
DOI:10.1002/bip.10521
Ribonucleoproteins (RNP) are involved in many essential processes in life. However, the roles of RNA and protein subunits in an RNP complex are often hard to dissect. In many RNP complexes, including the ribosome and the Group II introns, one main function of the protein subunits is to facilitate RNA folding. However, in other systems, the protein subunits may perform additional functions, and can affect the biological activities of the RNP complexes. In this review, we use ribonuclease P (RNase P) as an example to illustrate how the protein subunit of this RNP affects different aspects of catalysis. RNase P plays an essential role in the processing of the precursor to transfer RNA (pre-tRNA) and is found in all three domains of life. While every cell has an RNase P (ribonuclease P) enzyme, only the bacterial and some of the archaeal RNase P RNAs (RNA component of RNase P) are active in vitro in the absence of the RNase P protein. RNase P is a remarkable enzyme in the fact that it has a conserved catalytic core composed of RNA around which a diverse array of protein(s) interact to create the RNase P holoenzyme. This combination of highly conserved RNA and altered protein components is a puzzle that allows the dissection of the functional roles of protein subunits in these RNP complexes. © 2003 Wiley Periodicals, Inc. Biopolymers 73: 79–89, 2004
Co-reporter:Craig P. McClure, Kristin M. Rusche, Katrina Peariso, Jane E. Jackman, Carol A. Fierke, James E. Penner-Hahn
Journal of Inorganic Biochemistry 2003 Volume 94(1–2) pp:78-85
Publication Date(Web):1 February 2003
DOI:10.1016/S0162-0134(02)00611-6
Extended X-ray absorption fine structure (EXAFS) spectroscopy has been used to determine the structure of the Zn(II) sites in UDP-(3-O-acyl)-N-acetylglucosamine deacetylase (LpxC) from Aquifex aeolicus and Pseudomonas aeruginosa. The active site Zn(II) is four coordinate, with exclusively low-Z (nitrogen and oxygen) ligation in both enzymes. The amplitude of the outer-shell scattering from the histidine ligands is best fit using two histidine ligands, suggesting a ZnO2(His)2 site, where O most likely represents a conserved aspartate and a solvent molecule. The same structure was found for Co(II)-substituted A. aeolicus LpxC, although in this case it is possible that the coordination sphere may expand to include a fifth low-Z ligand. EXAFS data were also measured for the Escherichia coli LpxC enzyme. When a single Co(II) is substituted for Zn(II) in the active site of E. coli LpxC, EXAFS data show the same ligand environment as is found for the P. aeruginosa and A. aeolicus enzymes. However, the EXAFS data for E. coli LpxC with two zinc ions bound per protein, with the second Zn(II) acting as an inhibitory metal, demonstrates that the inhibitory metal is bound to at least two high-Z (sulfur, presumably thiolate, or chlorine) ligands. Results of the outer-shell scattering analysis, combined with previous studies of the LpxC enzyme, indicate a novel zinc binding motif not found in any previously studied zinc metalloproteins.
Co-reporter:Jennifer S Griffiths, Nathan J Wymer, Eugenia Njolito, S Niranjanakumari, Carol A Fierke, Eric J Toone
Bioorganic & Medicinal Chemistry 2002 Volume 10(Issue 3) pp:545-550
Publication Date(Web):March 2002
DOI:10.1016/S0968-0896(01)00307-8
The Thermotoga maritima aldolase gene has been cloned into a T7 expression vector and overexpressed in Escherichia coli. The preparation yields 470 UL−1 of enzyme at a specific activity of 9.4 U mg−1. During retroaldol cleavage of KDPG, the enzyme shows a kcat that decreases with decreasing temperature. A more than offsetting decrease in Km yields an enzyme that is more efficient at 40 °C than at 70 °C. The substrate specificity of the enzyme was evaluated in the synthetic direction with a range of aldehyde substrates. Although the protein shows considerable structural homology to KDPG aldolases from mesophilic sources, significant differences in substrate specificity exist. A preparative scale reaction between 2-pyridine carboxaldehyde and pyruvate provided product of the same absolute configuration as mesophilic enzymes, but with diminished stereoselectivity.Graphic
Co-reporter:James L. Hougland, Katherine A. Hicks, Heather L. Hartman, Rebekah A. Kelly, ... Carol A. Fierke
Journal of Molecular Biology (8 January 2010) Volume 395(Issue 1) pp:176-190
Publication Date(Web):8 January 2010
DOI:10.1016/j.jmb.2009.10.038
Prenylation is a posttranslational modification essential for the proper localization and function of many proteins. Farnesylation, the attachment of a 15-carbon farnesyl group near the C-terminus of protein substrates, is catalyzed by protein farnesyltransferase (FTase). Farnesylation has received significant interest as a target for pharmaceutical development, and farnesyltransferase inhibitors are in clinical trials as cancer therapeutics. However, as the total complement of prenylated proteins is unknown, the FTase substrates responsible for farnesyltransferase inhibitor efficacy are not yet understood. Identifying novel prenylated proteins within the human proteome constitutes an important step towards understanding prenylation-dependent cellular processes. Based on sequence preferences for FTase derived from analysis of known farnesylated proteins, we selected and screened a library of small peptides representing the C-termini of 213 human proteins for activity with FTase. We identified 77 novel FTase substrates that exhibit multiple-turnover (MTO) reactivity within this library; our library also contained 85 peptides that can be farnesylated by FTase only under single-turnover (STO) conditions. Based on these results, a second library was designed that yielded an additional 29 novel MTO FTase substrates and 45 STO substrates. The two classes of substrates exhibit different specificity requirements. Efficient MTO reactivity correlates with the presence of a nonpolar amino acid at the a2 position and a Phe, Met, or Gln at the terminal X residue, consistent with the proposed Ca1a2X sequence model. In contrast, the sequences of the STO substrates vary significantly more at both the a2 and the X residues and are not well described by current farnesylation algorithms. These results improve the definition of prenyltransferase substrate specificity, test the efficacy of substrate algorithms, and provide valuable information about therapeutic targets. Finally, these data illuminate the potential for in vivo regulation of prenylation through modulation of STO versus MTO peptide reactivity with FTase.
Co-reporter:John Hsieh, Kristin S. Koutmou, David Rueda, Markos Koutmos, ... Carol A. Fierke
Journal of Molecular Biology (2 July 2010) Volume 400(Issue 1) pp:38-51
Publication Date(Web):2 July 2010
DOI:10.1016/j.jmb.2010.04.050
Metal ions interact with RNA to enhance folding, stabilize structure, and, in some cases, facilitate catalysis. Assigning functional roles to specifically bound metal ions presents a major challenge in analyzing the catalytic mechanisms of ribozymes. Bacillus subtilis ribonuclease P (RNase P), composed of a catalytically active RNA subunit (PRNA) and a small protein subunit (P protein), catalyzes the 5′-end maturation of precursor tRNAs (pre-tRNAs). Inner-sphere coordination of divalent metal ions to PRNA is essential for catalytic activity but not for the formation of the RNase P·pre-tRNA (enzyme–substrate, ES) complex. Previous studies have demonstrated that this ES complex undergoes an essential conformational change (to the ES⁎ conformer) before the cleavage step. Here, we show that the ES⁎ conformer is stabilized by a high-affinity divalent cation capable of inner-sphere coordination, such as Ca(II) or Mg(II). Additionally, a second, lower-affinity Mg(II) activates cleavage catalyzed by RNase P. Structural changes that occur upon binding Ca(II) to the ES complex were determined by time-resolved Förster resonance energy transfer measurements of the distances between donor–acceptor fluorophores introduced at specific locations on the P protein and pre-tRNA 5′ leader. These data demonstrate that the 5′ leader of pre-tRNA moves 4 to 6 Å closer to the PRNA·P protein interface during the ES-to-ES⁎ transition and suggest that the metal-dependent conformational change reorganizes the bound substrate in the active site to form a catalytically competent ES⁎ complex.
Co-reporter:Kristin S. Koutmou, Nathan H. Zahler, Jeffrey C. Kurz, Frank E. Campbell, ... Carol A. Fierke
Journal of Molecular Biology (12 February 2010) Volume 396(Issue 1) pp:195-208
Publication Date(Web):12 February 2010
DOI:10.1016/j.jmb.2009.11.039
Bacterial ribonuclease P (RNase P) catalyzes the cleavage of 5′ leader sequences from precursor tRNAs (pre-tRNAs). Previously, all known substrate nucleotide specificities in this system are derived from RNA–RNA interactions with the RNase P RNA subunit. Here, we demonstrate that pre-tRNA binding affinities for Bacillus subtilis and Escherichia coli RNase P are enhanced by sequence-specific contacts between the fourth pre-tRNA nucleotide on the 5′ side of the cleavage site (N(− 4)) and the RNase P protein (P protein) subunit. B. subtilis RNase P has a higher affinity for pre-tRNA with adenosine at N(− 4), and this binding preference is amplified at physiological divalent ion concentrations. Measurements of pre-tRNA-containing adenosine analogs at N(− 4) indicate that specificity arises from a combination of hydrogen bonding to the N6 exocyclic amine of adenosine and steric exclusion of the N2 amine of guanosine. Mutagenesis of B. subtilis P protein indicates that F20 and Y34 contribute to selectivity at N(− 4). The hydroxyl group of Y34 enhances selectivity, likely by forming a hydrogen bond with the N(− 4) nucleotide. The sequence preference of E. coli RNase P is diminished, showing a weak preference for adenosine and cytosine at N(− 4), consistent with the substitution of Leu for Y34 in the E. coli P protein. This is the first identification of a sequence-specific contact between P protein and pre-tRNA that contributes to molecular recognition of RNase P. Additionally, sequence analyses reveal that a greater-than-expected fraction of pre-tRNAs from both E. coli and B. subtilis contains a nucleotide at N(− 4) that enhances RNase P affinity. This observation suggests that specificity at N(− 4) contributes to substrate recognition in vivo. Furthermore, bioinformatic analyses suggest that sequence-specific contacts between the protein subunit and the leader sequences of pre-tRNAs may be common in bacterial RNase P and may lead to species-specific substrate recognition.
.jpg)


![1,6-Heptadiene-3,5-dione,1,7-bis[4-(acetyloxy)-3-methoxyphenyl]-, (E,E)-](http://img.cochemist.com/ccimg/19700/19697-86-0.png)
![1,6-Heptadiene-3,5-dione,1,7-bis[4-(acetyloxy)-3-methoxyphenyl]-, (E,E)-](http://img.cochemist.com/ccimg/19700/19697-86-0_b.png)